
D.O. Edem
Department of Biochemistry, Faculty of Basic Medical Sciences, University of Uyo, P.M.B. 1017, Uyo, Akwa Ibom State, Nigeria
Asian Journal of Clinical Nutrition
Year: 2009 | Volume: 1 | Issue: 1 | Page No.: 65-82







ABSTRACT
This study provides a comprehensive review on the fat-soluble micronutrient vitamin A, changes in its status among vulnerable groups of different populations and the results of experience with different strategies/interventions designed to improve its status. Vitamin A can be derived from animal sources (preformed in liver, milk, eggs, fatty fish) or plant sources (in fruits and vegetables as provitamin A). The vitamin is required for normal growth and development, playing a part in reproduction, differentiation of cellular epithelium, regulation of cell division, genetic regulation and enhancement of immune responses. Retinol (its basic molecule) is metabolised into a number of biologically active retinoid compounds, such as retinal (active element of visual pigment) and retinoic acid, an intracellular messenger that modulates cell differentiation. Uneven distribution of natural sources of vitamin A, low dietary intake of animal products, the traditional technological transformation/processing of foods, low absorption and limited bioconversion of carotenoids may impact negatively on vitamin A activity. Vitamin A deficiency (VAD), a major public health nutritional problem in many developing countries, results in a range of developmental and pathological states with consequences ranging from potentially blinding xerophthalmia to increased risks of infection and mortality. All alternatives employed towards eradicating VAD (nutrition education, dietary diversification, dietary supplementation and food fortification) have provided a high return on investment in terms of disability-adjusted-life years gained. Key options for action, as well as important areas of research are being discussed. Control of VAD will lead to a substantial improvement in childhood survival and prevent the scandal of irreversible blindness. It is suggested that early introduction of children to abundant natural sources of vitamin A (like red palm oil-prepared meals) be encouraged.
PDF Abstract XML References Citation
How to cite this article
D.O. Edem, 2009. Vitamin A: A Review. Asian Journal of Clinical Nutrition, 1: 65-82.
DOI: 10.3923/ajcn.2009.65.82
URL: https://scialert.net/abstract/?doi=ajcn.2009.65.82
DOI: 10.3923/ajcn.2009.65.82
URL: https://scialert.net/abstract/?doi=ajcn.2009.65.82
INTRODUCTION
Vitamin A is an isoprenoid compound with a 6-membered ring and an 11-carbon side chain. The basic molecule of vitamin A is retinol (vitamin A alcohol), which is present in foods and tissues mainly as esters combined with long-chain fatty acids such as palmitic acid (Bates, 1995; Wongsiriroj and Blaner, 2004). Retinol (all trans form, vitamin A1) is the most biologically active and commonest form in mammalian tissues. It forms pale yellow crystals soluble in organic solvents and fats. It is easily destroyed by ultraviolet light, acids, oxygen and heat (Anosike, 1994). Vitamin A is found in plants as a provitamin called β-carotene (the most abundant carotenoid which can be converted to vitamin A by an oxygenase present in the intestine). Carotenoids are abundantly distributed in plants and plant foods (Englberger, 1999; Rao, 2000) and are readily available to the human body from some usual diets (Kritchevsky, 2000). The carotenoids (in addition to vitamin E, ascorbic acid, enzymes and proteins) are members of the biological antioxidant network (McLaren and Frigg, 2001) converting highly reactive radicals (`OH) and free fatty peroxy radicals to less active species, thus protecting against unwanted oxidative events. The structures of vitamin A and some of its metabolites are shown in Table 1, while the conversion of β-carotene (all trans) to vitamin A (retinal) is shown in Fig. 1.
| Table 1: | Principal members of the group of vitamin A |
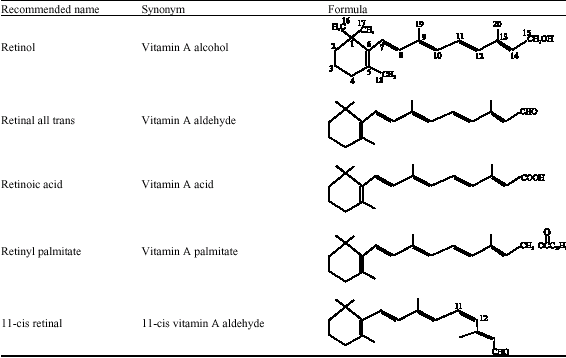 | |
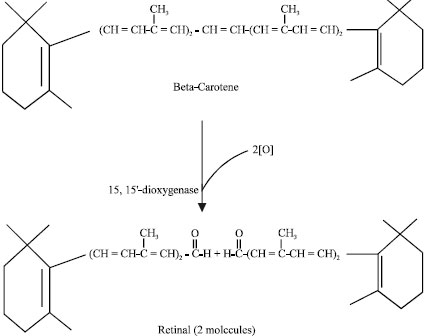 | |
| Fig. 1: | Conversion of β-carotene to retinal |
Biochemical Action of Vitamin A
Vitamin A is required for normal development and growth (Rondo, 2001; McCaffery et al., 2003). It plays a role in many metabolic processes in many cells, but its best-known function is in the eye where it plays an important role in the visual process (Saari, 1999). Vitamin A plays a part in reproduction [synthesis of sexual steroids, spermatogenesis, conception and placenta formation] (Gudas et al., 1994; Alais and Linden, 1999; Chung et al., 2005), differentiation of cellular epithelium, regulation of cell division, bone remodelling, genetic regulation, enhancement of immune response (Olson, 1994 a, b; Semba, 1994; Bates, 1995; Zachman, 1995; Thurnham et al., 2000; Serghides and Kain, 2002) and in all stages of lung development (Zachman, 1995; Sempertegui, 2001). Retinol (vitamin A1) is metabolised into a number of biologically active retinoid compounds (Morris-Kay, 1992; Blomhoff, 1994; Sporn et al., 1994; Mangelsdorf et al., 1995). The retinoids are important signalling molecules in embryonic development and in cellular differentiation throughout adult life, for example in epithelial and lymphoid differentiation (Morris-Kay, 1992; Blomhoff, 1994; Sporn et al., 1994). The significance of retinoid signals is reflected in the teratogenic effects of retinoids in humans and other animals (Morris-Kay, 1992; Rothman et al., 1995; Wiegand et al., 1998; McCaffery et al., 2003) and in the successful use of retinoids as treatments for some skin diseases and some forms of cancer (Blomhoff, 1994; Lamm et al., 1994; Sporn et al., 1994). Two important active metabolites are retinoic acid (an intracellular messenger that modulates cell differentiation) and retinal (or retinaldehyde), the active element of visual pigment.
Rhodopsin (visual purple), the light-sensitive pigment (chromoprotein) in the rod cells of the retina consist of the protein called opsin combined with 11-cis retinal (the oxidized or aldehyde form of retinol). It is used for vision in dim light. During this process, light isomerises the 11-cis retinal. Because the conformation of all trans retinal does not fit into the binding site for 11-cis retinal, the rhodopsin molecule becomes unstable and undergoes a series of conformational changes or molecular modifications followed by the hydrolysis of the Schiff`s base linkage (of rhodopsin) between all-trans retinal and opsin to yield opsin and trans-retinal (Anosike, 1994). This dissociation gives rise to visual excitement and the sensation of light perceived during the period of conformational changes (in the rhodopsin molecule) is transmitted by the optic nerve to the visual centres of the brain which translate as images. A nerve impulse is initiated during the rapid changes of protein conformation. The dissociation of rhodopsin changes its colour from purple (red) to yellow and is thus said to be bleached. Trans retinal may isomerise directly to 11-cis retinal, which recombines with opsin to form rhodopsin; alternatively it may first be reduced to retinol by retinal reductase followed by isomerization of the retinal and reoxidation to 11-cis retinal. The visual cycle of vitamin A is shown in Fig. 2.
Some retinal is continually lost from the cells of the retina, so that the continuation of the visual cycle depends upon a continual replacement from the diet through the blood supply to the retina (Anosike, 1994). Similar visual cyclic processes occur in both the rod and the cone cells. Each cone cell contains 3 different colour-sensitive pigments (iodopsins), which absorb light of wavelengths 430, 540 and 575 nm, respectively. These iodopsins contain 11-cis retinal but their opsins (cone-type opsins) are different.
Retinoic acid (another important metabolite of vitamin A) is formed when an enzyme NADH-dependent aldehyde dehydrogenase acts on retinal bound to cellular retinal binding protein CRBP. This compound modulates cell differentiation (Bates, 1995; Wang et al., 2005). Retinoic acid (RA) binds to its own cellular retinoic acid receptors (RAR), which are ligand-activated DNA binding proteins that belong to the steroid-thyroid-retinoid super family of nuclear receptors and their activators (Kasner et al., 1994; Nilsson et al., 1997; Rowe, 1997; Blaner, 1998; McLaren and Frigg, 2001; Balmer and Blomhoff, 2002). RAR then interacts with response elements (RARE) to affect a target gene. Thus RA potentially can regulate the expression and products of many different genes in many cell types. At a molecular level, more than 300 genes have already been identified whose expressions are influenced by vitamin A status (Murray et al., 2000). All stages of lung development (differentiation, growth and health) are potential targets of vitamin A influence (Zachman, 1995).
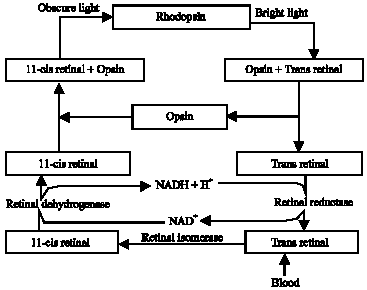 | |
| Fig. 2: | Visual cycle of vitamin A |
Retinoids (including RA) exert important effects on immune functions including favourable effect on responses to infections, differentiation, stimulation of phagocytosis and modulation of cytokines and eicosanoids (Bates, 1995; D-Souza and D-Souza, 2002; Serghides and Kain, 2002; Ihedioha et al., 2003; Wilson et al., 2003). Retinoic acid and β-carotene are inducers of gap-junctional or cell-cell communication, exerting growth control over cancer-initiated cells and preventing their malignant transformation (Van Poppel, 1993). Gap junctions transfer growth regulatory signals from health cells to adjacent cancer-initiated cells preventing their progression to the malignant state.
When cells are transformed into cancer cells, cell differentiation is suppressed. The carotenoids and retinoids may prevent harmful DNA damage induced by free radicals in the early stages of carcinogenesis and may counteract the effect of free-radical generating tumour promoters thus inhibiting both the initiation and promotion phases of carcinogenesis (Germann, 1994; Gerster, 1993). These compounds enhance the synthesis of the protein connexin which is responsible for gap junction communication between cells in culture (Bates, 1995).
Vitamin A has been shown to potentiate the antibody response to a variety of antigens, including rotavirus, E. coli and cholera toxin (Ahmed et al., 1991; Friedman et al., 1991; Wiedermann et al., 1993) as well as T-cell function (Coutsoudis et al., 1992; Semba, 1994).
Vitamin A decreases childhood mortality by reducing the risk of developing persistent diarrhoea (Bhandari et al., 1997; Rahman et al., 2001). This is true because the correction of sub clinical vitamin A deficiency results in a rapid and effective repair of the intestinal epithelium following an acute infection, owing to the role of vitamin A in regulating cell division (Semba, 1994) or enhancing immune response. Persistent diarrhoea accounts for 23 to 62% of diarrhoea-associated deaths in developing countries (Black, 1993).
Vitamin A and its precursor β-carotene, when added to iron-fortified grain products (e.g., rice, wheat or corn), appear to increase iron absorption by preventing the inhibitory effect of phytates on the absorption of non-heme iron from grain products. Vitamin A or β-carotene may form a complex with iron, keeping it soluble in the intestinal lumen (Garcia-Casal et al., 1998).
Interaction of Vitamin A with Other Nutrients
In the course of its metabolism, vitamin A interacts with other nutrients. These interactions influence absorption, metabolism, or excretion of the vitamin and/or these nutrients. The interactions of vitamin A with vitamin E, iron, zinc, copper, lipids and proteins are discussed below.
Vitamin E protects vitamin A from intracellular and intraluminal oxidation. It plays an important part in co-operating with vitamin A in the photoreceptors of the retinal cells where it protects retinaldehyde from oxidation (McLaren and Frigg, 2001). Retinyl esters are stored in the liver and other tissues in tocopherol-rich lipid globules (Olson, 1994b).
Iron (Fe) is an element necessary for several metabolic functions. The role of Fe is mostly confined to the processes of cellular respiration, where it plays a fundamental role in the transport of oxygen. Iron is a component of haemoglobin, myoglobin and cytochrome, as well as the enzymes catalase and peroxidase. During Fe deficiency, there is impaired mobilization of liver retinol (Strube et al., 2002). According to Muslimatum (2001) and Schmidt (2001), vitamin A plays a role in the activation of haematopoiesis by improving utilization of Fe for haemopoiesis and liberation of iron from iron stores. Retinol and/or retinoic acid are required for the synthesis of the iron transport protein transferrin (Devlin, 2006). Suharno et al. (1993) reported that supplementation with Fe and vitamin A can virtually eliminate nutritional anaemia (Vitamin A + Fe = Higher haemoglobin). Iron is required by two enzymes in the metabolic pathway for the conversion of tryptophan (try) to nicotinic acid nucleotides: (i) the heme enzyme try pyrrolase (tryptophan -2, 3-dioxygenase) and (ii) 3-hydroxyanthranilic acid dioxygenase [Fe++ dependent] (Van Eys, 1991). Iron deficiency results in reduction in heme cofactor and hence decreases of the rate-limiting enzyme try dioxygenase (in the conversion of try to its end products). Nicotinic acid nucleotides (e.g., NAD) are required by enzymes involved in the interconversions of vitamin A substances in the visual process. Thus, Fe deficiency may adversely affect the visual process.
Zinc (Zn) is necessary to maintain normal concentrations of vitamin A in plasma. Zinc deficiency impairs efficiency of β-carotene conversion to vitamin A. It is also required for normal mobilization of vitamin A from liver (Suharno et al., 1993). The synthesis of retinol binding protein (RBP) (responsible for transport of vitamin A) takes place in the presence of Zn (McLaren and Frigg, 2001). Some enzymes in the metabolism of vitamin A are metalloenzymes in which Zn forms a prosthetic group. For instance, it has been demonstrated that Zn deficiency diminishes the activity of retinal reductase (which converts retinal to retinol) and increases the activity of retinol oxidase [which converts retinol to retinoic acid] (Zachman, 1995; Boron et al., 1988). The lymphatic output of retinol is decreased by low zinc intake (Noh Sang and Koo Sung, 2003).
Copper ions are required by tryptophan dioxygenase (Van Eys, 1991) the rate-limiting enzyme in the pathway for the conversion of tryptophan to nicotinic acid nucleotides. NAD is important in vitamin A metabolism.
The absorption of vitamin A and its precursors is conditioned by fats (Lietz et al., 2001; Hedren et al., 2002; Ribaya-Mercado, 2002; Mulokozi et al., 2004). After ingestion, the products of dietary lipid digestion (mono, di and triglycerides, fatty acids and phospholipids) interact with bile salts and cholesterol to form aggregates. The latter solubilize vitamin A and β-carotene. This results in emulsified particles, which diffuse in the glycoprotein brush border of the microvilli of the intestine where they are absorbed (Olson, 1994a, b).
There is a correlation between VAD and protein malnutrition. The transport of retinol across different organs is made possible by retinol-binding protein (RBP). It is possible that protein deficiency, which can lead to decreased synthesis of RBP may reduce the levels of vitamin A in the blood. Adequate protein (and Zn) intake assist maintenance of vitamin A status (McLaren and Frigg, 2001). Malnourished children have lower serum retinol levels than those well nourished (Oso et al., 2003).
Dietary Sources of Vitamin A
Vitamin A (preformed) occurs mainly in animal products such as milk, liver, meat, butter, egg yolk, cod liver oil and in animal fat. The richest food source is liver, with other animal and fish sources providing substantial amounts of the preformed vitamin (Bates, 1995; Roos et al., 2003). All pigmented (particularly orange and yellow) vegetables and fruits, leafy green vegetables and yellow corn supply provitamin A in the diet. All the carotenes that are abundant in green plants, certain fruits (like pawpaw, mangoes, oil palm fruits, pumpkins), carrots, sweet potatoes and spinach have provitamin A activity (Anosike, 1994; Scheuring et al., 1996; Shamin et al., 2003). Most vertebrates (except carnivores, which depend on vitamin A in the liver of animals they feed on) convert the carotenes to vitamin A by enzymatic reactions in the intestinal mucosa and liver (Whitney et al., 1990). In rats, absorbed β-carotene is almost completely converted and the liver is the main storage site of vitamin A (Van Vliet et al., 1996). Beta-carotene is oxidatively cleaved (by β-carotene-15, 15`- dioxygenase) into two molecules of retinal (α and γ-carotene each yield one molecule of vitamin A), which are then reduced to all-trans retinol to be esterified (by lecithin: retinol acyltransferase LRAT) with a long-chain fatty acid mainly palmitate. After absorption, retinol is transported via the chylomicrons to the liver, where it is either stored as retinyl esters, or re-exported into the plasma as the free alcohol in combination with RBP, from which it is delivered to critical tissue sites. Retinol is removed from the plasma by the cells of the retina and oxidized to all-trans retinal. It is reported that one retinol equivalent (RE) is provided by 1 μg of all trans retinol or by 6 μg all-trans β-carotene or 12 μg of other provitamin A carotenoids (NRC, 1989; Brody, 1999). (1RE = 1 μg retinol = 0.0035 μmol retinol; 1 μmol retinol = 286 μg retinol; 1 IU vitamin A = 0.3 μg retinol). The recommended daily allowance for vitamin A is 350-1300 RE (Murray et al., 2000).
A retinol concentration (in a healthy person of 0.07 μmol g-1 (or 20 μg g-1)) wet weight of liver seems to fulfil all physiological needs and to provide an adequate reserve for approximately a three-month period. In malnourished children, the normally large amounts of vitamin stored in the liver are virtually exhausted to levels below 15 μg g-1 fresh liver tissues and this can lead to a deficiency of the vitamin.
Deficiency of Vitamin A
Vitamin A deficiency (VAD) is a level of depletion of total body stores of retinol and of its active metabolites such that normal physiological function is impaired. It may also be considered as the presence of low serum levels of vitamin. A, which may result from a prolonged deficient dietary intake, depletion of total body stores of retinol (and of its active metabolites), such that normal physiological function is impaired (Northrop-Clewes, 2001; West, 2002). Vitamin A deficiency is caused by insufficient intake of foods rich in vitamin A or provitamin A carotenoids to meet the requirements for growth, infection, pregnancy and lactation (WHO, 1996).
Vitamin A (retinol) is essential to mammals. Its deficiency can result in a range of developmental and pathological states (such as heart, lung and eye defects), including blindness (Morriss-Kay, 1992; Blomhoff, 1994; Sporn et al., 1994). The deficiency of vitamin A can be indicated by serum (or breast milk) concentrations of retinol, retinol-binding protein and relative dose response (RDR) measurements. Deficiency is often associated with protein-energy malnutrition, parasitic infestations and diarrhoea disease (Blaauw, 1999; Grubesic, 2004). The functional indicators of VAD include papillary dark adaptation (PDA), conjunctival impression cytology (CIC) and xerophthalmia.
The earliest clinical sign of VAD is impaired vision in dim light. The deficiency of vitamin A causes night blindness (nyctalopia) because of the inability to regenerate rhodopsin in the rod cells of the retina (Kasner et al., 1994). VAD is among the most widespread and serious nutritional disorders that affect mankind (Humphrey et al., 1992; Jorymar et al., 2007). Worldwide, an estimated 2.8 million young children 0-4 years of age are clinically affected by VAD and 251 million more are severely or moderately sub clinically deficient. Thus at least 254 million pre-school children are at risk in terms of their health and survival (Humphrey et al., 1992; WHO, 1995) and this has accounted for more than 1 million deaths each year. Xerophthalmia (keratinisation of ocular tissues), which is among the oldest recorded manifestations of VAD known to man, is a major cause of blindness in childhood (Singh and West Jr., 2001; Dada, 2002). It also predisposes to increased diarrhoea and respiratory morbidity (Sommer et al., 1984; Feachem, 1987; Gujral et al., 1993). Vitamin A deficiency exists in more than 60 countries and affects more than 7.2 million pregnant women (West, 2002). About 250,000 children of preschool age become blind each year (WHO, 1992). Today VAD and its ocular manifestations are largely limited to the developing countries, particularly those in Africa, Asia (which houses more than half of the world`s population) and Western Pacific (Udomkesmalee, 1992; Jorymar et al., 2007). Mortality primarily from infectious disease has been identified as an important accompaniment of xerophthalmia and has now proved to be responsive to massive intermittent vitamin A dosage schedules that were originally introduced to combat blindness (Sommer et al., 1986; Glasziou and Mackerras, 1993), thus raising the possibility that vitamin A influences specific elements of the immune system (Coutsoudis et al., 1992; Ross and Hammerling, 1994; Bates, 1995; Zhao and Ross, 1995).
The clinical signs of VAD appear when the average intake of the vitamin is very low, which corresponds to negligible liver reserves. Plasma retinol concentrations of less than 10 μg dL-1 are usually associated with Bitot`s spots and other signs of deficiency or can be found in a vitamin A-sufficient child often plagued with infections.
Consequences of Vitamin A Deficiency
Vitamin A deficiency is the leading cause of preventable blindness in children Earliest indications of deficiency are usually night blindness and Bitot`s spots (pathological changes in the eye). Some stages of ocular involvement (e.g., night blindness, conjunctival xerosis, Bitot`s spots and corneal xerosis) are entirely reversible with appropriate treatment, while later stages (e.g., corneal ulceration/keratomalacia, corneal scar and xerophthalmic fundus) result in permanent eye damage. Subclinical VAD (without ocular symptoms) has been associated with an immune response that results in an increased risk of fatality in measles (Rosales, 2002). Immediate treatment with retinol can save lives in such cases.
Vitamin A deficiency causes replacement of mucus-secreting epithelium in the trachea and bronchi. The differentiation of various pulmonary cells studied in culture requires retinoids (Chytil, 1992). Pulmonary morbidity is frequently associated with human VAD (Fawzi et al., 1993). Some persons speculate that vitamin A may have a role in the prevention and / or repair of lung injury in human premature new born (Shenai, 1994; Zachman, 1995; Spears et al., 2004).
Epithelial damage resulting from VAD would favour invasion and colonization by bacteria, when lesions of the eye are apparent and plasma levels of vitamin A are very low -<1.05 mmol L-1 (Murray et al., 2000; West, 2002). The resulting infection may play a role in the aetiology of nutritional anaemia; it depresses the synthesis of retinol binding protein and transferrin in addition to causing an increase in endothelial permeability (Thurnham, 1993).
Vitamin A deficiency in the rat causes desquamation of the germinal cells of the seminiferous tubules and resorption of the foetus (Huang et al., 1985; Gudas et al., 1994). This highlights the importance of vitamin A in reproduction. Parke and Ioannides (1981) had reported that VAD decreases liver cytochrome P-450 and mixed function oxidases (MFO) enzymes which explains it effects on the induction of certain enzymes for the detoxification of xenobiotics in animals e.g., aflatoxin B1 O-demethylase, aflatoxin B1 hydoxylase, ethylmorphine N-demethylase and aniline hydroxylase (Halvadar, 1991).
The inflammatory response in the clinical manifestations of inflammatory reactions may be increased by high production of interferon-γ during VAD (Feuerer et al., 2006).
Reasons for VAD
Vitamin A activity can be derived from animal sources (preformed vitamin A from milk, liver, fatty fish, meat, eggs) or from fruits and vegetables (β-carotene and other carotenoids = provitamin A). In spite of the large diversity of foods rich in vitamin A or provitamin A, these are not within the reach of all social groups (Gouado and Ejoh, 2005) The existing supply of vitamin A from natural sources, if distributed evenly among the population, would be sufficient to meet the needs of the people (FAO, 1992). Availability of vitamin A may be satisfactory, but intake is inadequate.
Many people in developing countries have low dietary intakes of animal products rich in preformed vitamin A. This may be because of the cost or for cultural reasons. Cultural traits may outweigh nutritional knowledge and buying power. Getting people to change their dietary habits and traditions is no easy matter. Man is wish-selective rather than need-selective (Brunken and Flores, 1994).
In most tropical countries of the world where VAD occurs, there is usually an ample supply of carotenoid-rich foods to provide adequate amounts of the vitamin. The problem is that many people do not include these foods in their diet for reasons of poverty and ignorance. The least expensive foods containing vitamin A are generally not very popular among these populations. When climatic conditions limit agricultural production, availability of plant foods rich in provitamin A carotenoids will be restricted; such foods may be excluded from the diet for cultural and other reasons (Asonye, 2003).
Furthermore the traditional technological transformation and processing of fruits and vegetables (bleaching of oil, excessive cooking of leafy vegetables) may affect the tenure of vitamin A and β-carotene in foods like palm oil and vegetables (Cottrell, 1991; Gouado et al., 1998; Ajose et al., 2004; Mulokozi et al., 2004). The bioavailability of vitamin A from plants may be limited (lower) and more variable than previously thought (Blum, 1997). Low absorption and limited bioconversion of carotenoids may limit the vitamin A activity of carotenoids. The factors responsible for low absorption include: low fat content of diet, carotenoids trapped within structural matrix of plant cells, malabsorption (interference with intestinal absorption) due to low pool of bile acids and diarrhoea due to poor hygiene. Mucosal damage due to infectious disease, diarrhoea and intestinal parasites has been responsible for low bioconversion (Blum, 1997). Interference with blood transport or liver storage of the vitamin may also cause deficiencies.
The vitamin A status of children may be further compounded by infectious diseases, which rapidly deplete vitamin A stores by increasing retinol utilization and excretion. A single episode of infection can cause VAD (Mitra et al., 1998). Improving vitamin A status can impact positively on child survival (Ross, 1996).
Elimination (Control) of VAD
Vitamin A deficiency remains one of the major public health nutritional problems in many developing countries (Thatcher et al., 1998; Dada, 2002; Rosales, 2002; Lin, 2003; Singh and West Jr, 2004) with consequences ranging from potentially blinding xerophthalmia to increased risks of infection and mortality (WHO, 1992; Humphrey et al., 1996; Sommer et al., 1996; West, 2002). Four key strategies have been recommended (by international organisations active in micronutrient projects (viz., WHO, UNICEF and International Vitamin A Consultative Group) to eradicate VAD. These include nutrition education, dietary diversification, dietary supplementation and food fortification.
Nutrition education (which creates micronutrient awareness) is an essential part of any programme. It is aimed at improving practices related to consumption of vitamin A-rich sources. It has been used successfully to introduce home gardening and new agricultural production methods, which aim at increasing the availability of vitamin A-rich food sources, in addition to providing additional income used mainly for purchasing food (Scheuring et al., 1996; Vijayaraghavan et al., 1997; Tumwegamire et al., 2004) Food choices are often made based on availability, price, personal preferences, habits and cultural traditions/taboos. Dietary improvement (through educational and horticultural interventions) is the most logical sustainable strategy to prevent VAD. Promotion of market gardening is necessary for VAD elimination. Nutrition education (which seems to be an important medium-term measure for improving vitamin A status, especially of vulnerable groups in the community), can be carried out through schools and health centres i.e., can be instutionalized. This should be coupled with social marketing of vitamin A-rich foods, using a variety of means including mass communication. A previously neglected food source can be made popular through social marketing.
Eating a variety of foods (dietary diversification) is the natural way to obtain the nutrients needed for health. It requires people to change their eating habits. It may also involve the need to increase production, distribution, consumption and preservation of micronutrient crops and animal products. Agricultural research is trying to diversify the sources and improve the yields of foods rich in provitamin A. Genetic engineering has made possible the development of rice with high quality protein, improved bioavailability and provitamin A (Ye et al., 1997; Potrykus, 2001; Schiermeier, 2001; Chong, 2003; Potrykus, 2003).
Administration of dietary supplements is an effective way to rapidly correct existing deficiencies or avoid their development in high-risk populations (Rosales, 2002; Basu, 2003). It is mainly indicated as an emergency strategy. It has been successfully implemented (high dose vitamin A capsules) to reduce suffering in vitamin A-deficient children. To produce adequate liver stores of vitamin A at 6 months of age, it is recommended to give 200,000 IU to the mothers at delivery and 4 doses of 50000 IU to the infants (at birth and at 6,10,14 weeks) with immunization contacts. Such doses are safe and expected to reduce morbidity/mortality (Humphrey and Rice, 2000). Vitamin A supplementation reduced the prevalence of diarrhoea by 36% (Bhandari et al., 1994). Rukmini (1994) reported that Indian school children fed on supplementary snacks prepared with red palm oil (natural source of β-carotene, 375 mg g-1) for 60 days had significant increases in serum retinol levels as well as an increased retinol store suggesting the ready bioavailability of β-carotene.
Also, some studies carried out by Gouado (1994) revealed that a supplementation trial with red palm oil (RPO) carried out for 5 days on 34 human subjects living in Laroa Village in the Far-North province of Cameroun (a community which does not generally consume palm oil), helped to reduce the prevalence of vitamin A deficiency from 97.05 to 69.69%. Furthermore, significant increases in plasma vitamin A concentrations were observed when experimental animals were fed diets supplemented with RPO (Edem, 1999; Edem et al., 2002). Red palm oil from the palm fruit (Elaesis guineensis) is a source of oil-emulsified α- and β-carotene in abundant concentrations (Narasinga Rao, 2000; Ong and Goh, 2002). Periodic dosing with RPO (in Orissa province of Eastern India) produced the same degree of vitamin A status prophylaxis, as did the more traditional administration of 200,000 IU of retinyl palmitate (Mohapatra and Manorama, 1997). Thus the β-carotene of RPO may be regarded as the key to a safe and effective vitamin A supplement, fortificant and food-based solution all wrapped in one (Solomons, 1998; Solomons and Orozco, 2003; Spinnler, 2003). Vuong et al. (2002) reported on the potential of the gac fruit Momordica cochinchinensis (native to Vietnam and richer in β-carotene than RPO) for improving vitamin A and β-carotene in Vietnamese children.
Food fortification (addition of vitamins, minerals and trace elements to staple foods) has been practised in numerous industrialized countries with considerable success. It is an effective public health measure to correct nutritional deficiencies in whole population or specific segments at risk. It does not require the active participation of the consumer. Traditional food consumption patterns can be maintained. The costs are negligible compared with the medical and public health costs associated with malnutrition. Food fortification is a very safe way to supplement the diet of the population, because the amounts of the micronutrient used are many times below possibly critical levels. Fortification often supplies groups at high risk. The advantages of food fortification include the following: (a) It is affordable (b) It is effective (c) It brings a high return on investment (d) Behavioural changes are not required (e) Fortification is socially acceptable and politically attractive (f) High population coverage can be achieved (g) The risk of toxicity is negligible (h) It is sustainable. The prospects for vitamin A fortification of staple foods are good-to-excellent in many countries. Dietary staples are the underpinnings of cultural food ways. Everyone in the society would consume quantities of the dietary staple. A given nutrient should be added to a staple food to ensure that everyone in the society receives it. The enrichment of foods is one of the most promising ways to avoid VAD (Flores et al., 1994; Kraus et al., 1998; Dary and Mora, 2002). In Guatemala, Honduras, El Salvador, Bolivia, Brazil, Colombia and many countries in the Latin-American-Caribbean region, efforts to control VAD through fortification of sugar are producing encouraging results (Mora, 1995, 2003; Bay, 1997). In some African countries (e.g., Zambia), sugar and maize meal have been fortified with vitamin A. Two brands of cooking fat which command a large share of the Kenyan market are fortified with vitamin A. Fortification of margarine (cooking oil) with vitamin A is already mandatory in the Philippines (Mahensho, 1994; Capanzana and Barba, 2004) and South Africa (World Bank, 1994). There have been attempts to fortify cassava meals (Asonye, 2001). Maize meal (in Zimbabwe and Namibia), flour and bread (Zimbabwe) have been fortified with vitamin A (World Bank, 1994). Flores et al. (1994) reported that the relative dose response in 83 deprived children fed rice fortified with artificially produced rice kernels containing vitamin A was similar to that obtained after administration of vitamin A (Arovit) directly, thus indicating an acceptable bioavailability of vitamin A in the premix. Rice is the staple grain that may represent the staple for at least half of mankind (Surridge, 2004). Its quality can be enhanced genetically. An effort in genetic engineering has been applied to inserting into the rice (Oryza sativa) the genetic material that enables it to make β-carotene (Ye et al., 1997; Schiermeier, 2001). Fortification delivers low but continuous levels of vitamin A and therefore it is safe.
Selective Criteria for VAD Control Strategy
The choice of an appropriate strategy to eliminate VAD is not a simple yes-or-no decision. Usually a combination of activities is required. A right mix of interventions tailored to the local circumstances should be applied in achieving the objective of VAD control. An acute problem, with clinical symptoms of deficiency will require a solution that provides fast relief in a well-defined target group. In this case, supplementation will probably be the first choice. Supplementation is a short-term therapeutic intervention. To improve vitamin A intakes of the population in the medium term, fortification should be considered. Fortification may also be a stable long-term solution. Dietary diversification and nutrition education are long-term alternatives that may take many years to become effective. Dietary diversification must include a variety of vegetables and fruits as well as animal foods like milk and eggs (De Pee et al., 1998). Dietary improvement (achievable through educational and horticultural interventions) is the most logical and sustainable strategy to prevent VAD.
Estimates show that all alternatives provide a high return on investment in terms of disability-adjusted life-years gained. The cost of delivering adequate amounts of vitamin A to individuals at high risk of VAD is lowest with food fortification (less than US $1 per year) and highest with gardening project (US $2-4 per year) with the cost of supplementation between them (World Bank, 1994).
However, over employment of the above strategies may lead to vitamin A toxicity or hypervitaminosis A.
Hypervitaminosis A
An increasingly vitamin A-rich diet has its downside aspects, as can be seen in the issues of teratogenesis risk for fertile-age women (Rothman et al., 1995) and life-long retinyl ester intakes for bone health of the mature adult (Melhus et al., 1998). Intake of vitamin A in the calcium sources milk, dairy products and fish above the level of 1500 RE daily begins to produce sufficient demineralization (Melhus et al., 1998). Penniston et al. (2003) suggested a link between chronic excessive intakes and predisposition to hip fractures. High doses of some retinoids have been reported to have harmful effects on the developing heart (Colbert, 2002). With intakes of greater than 500 mg of preformed retinyl esters, as occurred when European Arctic explorers (short of food) consumed liver (of polar bear [6000 RE g-1], marine animals like seal or their sled dogs), a severe and often fatal acute hypervitaminosis can result (Rothman et al., 1995; Murray et al., 2000; McLaren and Frigg, 2001; Devlin, 2006). With self-medication or unwise prescription of 7500 RE (retinol equivalents) of preformed vitamin A, chronic hypervitaminosis with neurological and hepatic consequences can occur. Hypervitaminosis A is an excessive vitamin A status in which there are excessive concentrations of retinol in plasma and symptoms/signs of toxicity (McLaren and Frigg, 2001). Acute toxicity is recognized by symptoms that suggest an acute rise in intracranial pressure-nausea, vomiting and headache. Some of the subjects have usually been young children in a supplementation programme. The symptoms of chronic toxicity include headache, vomiting, diplopia, alopecia, dryness of mucous membranes, desquamation, bone and joint pain, liver damage, bulging fontanel in children, hemorrhages and coma. Rothman et al. (1995) have estimated that daily intake of 3000 RE of retinol is sufficient to produce teratogenesis. Phytol and phytanic acid have been suggested to be useful for the prevention of vitamin A teratogenicity, possibly by reducing the metabolic production of retinoic acid (Arnhold et al., 2002).
Considering the deleterious effects of high doses of preformed vitamin A, the utility of β-carotene (whose absorption is down-regulated at higher intakes) has merit. Precursors of vitamin A (carotenoids) are regulated homeostatically in their conversion to vitamin A and cannot produce hypervitaminosis A (Mathews-Roth, 1993). However, toxicity of vitamin A is rare from natural diets with possible exception of liver, which contains 3-5 mg/100 g. Furthermore, many of the symptoms of vitamin A toxicity regress within a week of withdrawal of the vitamin (McLaren and Frigg, 2001).
CONCLUSION
Vitamin A status is an important factor in children morbidity and mortality. Vitamin A interacts beneficially with the immune system by promoting optimum protective host immune responses. There is compelling evidence in favour of a critical role for vitamin A in maintaining host defences against pathogens (Anderson, 1995). Vitamin A reinforces immune surveillance as well as inhibits carcinogenesis. Its immunologic effect is primarily to enhance generation of cytotoxic T-lymphocytes as part of cell-mediated immunity (Zachman, 1995).
Vitamin A malnutrition is an important health problem in many parts of the world. Each of the four strategies recommended as possible solutions has its strengths and weakness that should be analysed in the context of cultural factors and economics to obtain the maximum impact. Experience gained in developing and industrialised countries has shown that food fortification is efficacious and safe. Economic studies by the World Bank and other institutions active in public health research confirm that it is a sustainable and cost-effective solution. Food fortification improves nutritional status without requiring the active participation of the consumer: it affects taste and appearance of foods only minimally. Suitable technologies for the fortification of flour, rice, sugar, vegetable oils and dairy products have been developed and are transferable to local food industries in developing countries.
Early introduction of children to abundant natural sources of vitamin A like red palm oil-prepared meals should be encouraged. The benefit is that these sources on their own do not lead to vitamin A toxicity though they are effective in improving vitamin A status of individuals. Control of VAD will lead to a substantive and lasting improvement in childhood survival as well as preventing the scandal of irreversible blindness due to malnutrition.
REFERENCES
- Ahmed, F., D.B. Jones and A.A. Jackson, 1991. Effect of vitamin A deficiency on the immune response of mice to epizootic diarrhoea of infant mice (EDIM) rotavirus infection in mice. Br. J. Nutr., 65: 478-785.
CrossRefPubMedDirect Link - Ajose, O.A., D.A. Adelekan and E.O. Ajewole, 2004. Vitamin A status of pregnant Nigerian women: Relationship to dietary habits and morbidity. Nutr. Health, 17: 325-333.
PubMedDirect Link - Arnhold, T., M.M.A. Elmazar and H. Nau, 2002. Prevention of vitamin A teratogenesis by phytol or phytanic acid results from reduced metabolism of retinol to the teratogenic metabolite, All-trans-retinoic acid. Toxicol. Sci., 66: 274-282.
CrossRefPubMedDirect Link - Asonye, C.C., 2001. Fortification of common Nigerian food-Cassava meals. Food Nutr. Bull., 22: 423-426.
Direct Link - Balmer, J.E. and R. Blomhoff, 2002. Gene expression regulation by retinoic acid. J. Lipid Res., 43: 1773-1808.
CrossRefDirect Link - Basu, S., B. Sengupta and P.K.R. Paladhi, 2003. Single megadose of vitamin A supplementation of Indian mothers and morbidity in breastfed young infants. Postgrad. Med. J., 79: 397-402.
CrossRefPubMedDirect Link - Bhandari, N., M.K. Bhan and S. Sazawal, 1994. Impact of massive doses of vitamin A given to preschool children with acute diarrhoea on subsequent respiratory and diarrhoeal morbidity. Br. Med. J., 309: 1404-1407.
PubMedDirect Link - Bhandari, N., R. Bahl , S. Sazawal and M.K. Bhan, 1997. Breast-feeding status alters the effect of vitamin A treatment during acute diarrhoea in children. J. Nutr., 127: 59-63.
Direct Link - Black, R.E., 1993. Persistent diarrhoea in children of developing countries. Pediatr. Infect. Dis. J., 12: 751-761.
PubMedDirect Link - Boron, B., J. Hupert and D.H. Barch, 1988. Effect of zinc deficiency on hepatic enzymes regulating vitamin A status. J. Nutr., 118: 995-1001.
PubMedDirect Link - Chong, M., 2003. Acceptance of golden rice in the Philippine rice bowl. Nature, 21: 971-972.
CrossRefDirect Link - Chung, S.S.W., X.Y. Wang and D.J. Wolgemuth, 2005. Male sterility in mice lacking retinoic acid receptor α involves specific abnormalities in spermiogenesis. Differentiation, 73: 188-198.
CrossRefDirect Link - Colbert, M.C., 2002. Retinoids and cardiovascular developmental defects. Cardiovasc. Toxicol., 2: 25-39.
CrossRefDirect Link - Cottrell, R.C., 1991. Introduction: Nutritional aspects of palm oil. Am. J. Clin. Nutr., 53: 989S-1009S.
CrossRefDirect Link - Coutsoudis, A., P. Kiepiela, H.M. Coovadia and M. Broughton, 1992. Vitamin A supplementation enhances specific IgG antibody levels and total lymphocyte numbers, while improving morbidity in measles. Pediatr. Infect. Dis. J., 11: 203-209.
PubMedDirect Link - Dary, O. and J.O. Mora, 2002. Food fortification to reduce vitamin A deficiency. International vitamin A consultative group recommendations. J. Nutr., 132: 2927-2933.
Direct Link - De Pee, S., C.E. West, D. Permaesih, S. Maruti, K.D. Muhilal and J.G. Hautvast, 1998. Orange fruit is more effective than are dark green leafy vegetables in increasing serum concentration of retinol and β-carotene in school children in Indonesia. Am. J. Clin. Nutr., 68: 1058-1067.
PubMedDirect Link - D-Souza, R.M. and R. D-Souza, 2002. Vitamin A for preventing secondary infection in children with measles-a systematic review. J. Trop. Pediatr., 48: 72-77.
CrossRefPubMedDirect Link - Edem, D.O., O.U. Eka and I.B. Umoh, 2002. Feeding of red palm oil-supplemented diets to rats may impact positively on vitamin A status. Int. J. Food Sci. Nutr., 53: 285-291.
CrossRefPubMedDirect Link - Fawzi, W.W., T.C. Chalmers, M.G. Herrera and F. Mosteller, 1993. Vitamin A supplementation and child mortality. A meta analysis. JAMA, 269: 898-903.
PubMed - Feuerer, M., K. Eulenburg, C. Loddenkemper, A. Hammann and J. Huehn, 2006. Self limitation of Th 1-mediated inflammation by interferon-γ. J. Immun., 176: 2857-2863.
PubMedDirect Link - Flores, H., M.B. Guerra, A.C.A. Cavalcanti, F.A.C.S. Campos, M.C.N.A. Azevedo and M.B.M. Silva, 1994. Bioavailability of vitamin A in a synthetic rice premix. J. Food Sci., 59: 371-372.
CrossRefDirect Link - Friedman, A., A. Meidovsky, G. Leitner and D. Sklan, 1991. Decreased resistance and immune response to Escherichia coli infection in chicks with low or high intakes of vitamin A. J. Nutr., 121: 395-400.
PubMedDirect Link - Garcia-Casal, M.N., M. Layrisse, L. Solano, M.A. Baron, F. Arguello, D. Llovera, J. Ramirez, I. Leets and E. Tropper, 1998. Vitamin A and β-carotene can improve nonheme iron absorption from rice, wheat and corn by humans. J. Nutr., 128: 646-650.
PubMedDirect Link - Gerster, H., 1993. Anticarcinogenic effect of common carotenoids. Int. J. Vit. Nutr. Res., 63: 93-121.
PubMedDirect Link - Glasziou, P.P.and D.E.M. Mackerras, 1993. Vitamin A supplementation in infectious diseases: A meta analysis. Br. Med. J., 306: 366-370.
PubMedDirect Link - Gujral, S., R. Abbi and T. Gopaldas, 1993. Vitamin A supplementation and morbidity in children. J. Trop. Pediatr., 39: 89-92.
Direct Link - Halvadar, P.V., 1991. Recurrent vitamin a deficiency and fatty liver. J. Trop. Pediatr., 37: 87-88.
PubMedDirect Link - Hedren, E., G. Mulokozi and U. Svanberg, 2002. In vitro accessibility of carotenes from leafy green vegetables cooked with sunflower oil or red palm oil. Int. J. Food Sci. Nutr., 53: 445-453.
PubMedDirect Link - Huang, H.S.F., G. Dyrenfurth, G.L. Gunsalus and C. Hemberee, 1985. Effect of vitamin a deficiency upon gonadotropin response to gonadotropin-releasing hormone. Biol. Reprod., 33: 1176-1187.
PubMedDirect Link - Humphrey, J.H., K.P. West and A. Sommer, 1992. Vitamin A deficiency and attributable mortality among under-5-year olds. WHO Bull., 70: 225-232.
Direct Link - Humphrey, J.H., T. Agoestina, L. Wu, A. Usmen, M. Nurachim, D. Subardja, S. Hidayat, J. Tielsch, K.P. West and A. Sommer, 1996. Impact of neonatal vitamin a supplementation on infant morbidity and mortality. J. Pediatr., 128: 489-496.
PubMedDirect Link - Humphrey, J.H. and A.L. Rice, 2000. Vitamin A supplementation of young infants. Lancet, 356: 422-424.
CrossRefPubMedDirect Link - Kraus, V.M., H. Delisle and N.W. Solomons, 1998. Fortified foods contribute one half of recommended vitamin A intake in poor Guatemalan toddlers. J. Nutr., 128: 860-864.
PubMedDirect Link - Kritchevsky, D., 2000. Impact of red palm oil on human nutrition and health. Food Nutr. Bull., 21: 182-188.
Direct Link - Lamm, D.L., D.R. Riggs, J.S. Shriver, P.F. Van Gilder, J.F. Rach and J.I. Dettaven, 1994. Megadose vitamin in bladder cancer: A double -blind clinical trial. J. Urol., 151: 21-26.
PubMedDirect Link - Lin, K.G., 2003. Update on the prevalence of malnutrition among children in Asia. Nepal Med. Coll. J., 5: 113-122.
PubMedDirect Link - Mathews-Roth, M.M., 1993. Carotenoids in erythropoietic porphyria and other photosensitivity diseases. Ann. NY Acad. Sci., 691: 127-138.
CrossRefPubMedDirect Link - Melhus, H., K. Michaelsson, A. Kindmark, R. Bergstrom and K. Holmberg et al., 1998. Excessive dietary intake of vitamin a is associated with reduced bone mineral density and increased risk for hip fracture. Ann. Intern. Med., 129: 770-778.
Direct Link - Mitra, A.K., J.O. Alvarez and C.B. Stephenson, 1998. Increased urinary retinol loss in children with severe infections. Lancet, 351: 1033-1034.
CrossRefPubMedDirect Link - Mohapatra, S. and R. Manorama, 1997. The protective effect of red palm oil in comparison with massive vitamin a dose in combating vitamin a deficiency in Orissa, India. Asia Pac. J. Clin. Nutr., 6: 246-250.
PubMedDirect Link - Mulokozi, G., E. Hedren and U. Svanberg, 2004. In vitro accessibility and intake of β-carotene from cooked green leafy vegetables and their estimated contribution to vitamin a requirements. Plant Food Hum. Nutr., 59: 1-9.
CrossRefDirect Link - Nilsson, A., G. Trøen, L.B. Petersen, S. Reppe, K.R. Norum and R. Blomhoff, 1997. Retinyl ester storage is altered in liver stellate cells and in HL60 cells transfected with cellular retinol-binding protein type I. Int. J. Biochem. Cell Biol., 29: 381-389.
CrossRef - Noh Sang, K. and I. Koo Sung, 2003. Low zinc intake decreases the lymphatic output of retinol in rats infused intraduodenally with β-carotene. J. Nutr. Biochem., 14: 147-153.
CrossRefDirect Link - National Rearch Council NRC, 1989. Recommended Dietary Allowances. 10th Edn., National Academy Press, Washington, DC. USA., pp: 284.
Direct Link - Ong, A.S.H. and S.H. Goh, 2002. Palm oil: A healthful and cost-effective dietary component. Food Nutr. Bull., 23: 11-22.
CrossRefPubMedDirect Link - Oso, O.O., P.O. Abiodun, O.O. Omotade and D. Oyewole, 2003. Vitamin a status and nutritional intake of carotenoids of pre-school children in Ijaye-orile community in Nigeria. J. Trop. Pediatr., 49: 42-47.
CrossRefPubMedDirect Link - Parke, D.V. and C. Ioannides, 1981. The role of nutrition in toxicology. Ann. Rev. Nutr., 1: 207-234.
CrossRefDirect Link - Potrykus, I., 2003. Nutritionally enhanced rice to combat malnutrition disorders. Nutr. Rev., 61: 101-104.
CrossRefPubMedDirect Link - Rahman, M.M, S.H. Vermund, M.F. Wahed, C.J. Fuchs, A.H. Baqui and J.O. Alvarez, 2001. Simultaneous zinc and vitamin a supplementation in bangladeshi children: Randomised double-blind controlled trial. Br. Med. J., 323: 314-318.
CrossRefDirect Link - Roo, B.S.N., 2000. Potential use of red palm oil in combating vitamin A deficiency in India. Food Nutr. Bull., 21: 202-211.
CrossRefDirect Link - Ribaya-Mercado, J.D., 2002. Influence of dietary fat on β-carotene absorption and bioconversion into vitamin A. Nutr. Rev., 60: 104-110.
CrossRefDirect Link - Rondo, P.H.C., 2001. Vitamin a and neonatal anthropometry. J. Trop. Pediatr., 47: 307-310.
CrossRefPubMedDirect Link - Roos, N., M. Islam and S.H. Thilsted, 2003. Small fish is an important dietary source of vitamin a and calcium in rural Bangladesh. Int. J. Food Sci. Nutr., 4: 329-339.
CrossRefPubMedDirect Link - Rosales, F.J., 2002. Vitamin a supplementation of vitamin a deficient measles patients lowers the risk of measles-related pneumonia in Zambian children. J. Nutr., 132: 3700-3703.
Direct Link - Ross, A.C., 1993. Cellular metabolism and activation of retinoids: Roles of cellular retinol-binding proteins. FASEB J., 7: 317-327.
PubMedDirect Link - Rothman, K.J., L.L. Moore, M.R. Singer, U.S. Nguyen, S. Mannino and A. Milunsky, 1995. Teratogenicity of high vitamin a intake. New Eng. J. Med., 333: 1369-1373.
Direct Link - Rukmini, C., 1994. Red palm oil to combat vitamin A deficiency in developing countries. Food Nutr. Bul., 15: 126-129.
Direct Link - Schiermeier, Q., 2001. Designer rice to combat diet deficiencies makes it debut. Nature, 409: 551-551.
Direct Link - Semba, R.D., 1994. Vitamin A, immunity and infection. Clin. Infect. Dis., 19: 489-499.
PubMedDirect Link - Sempertegui, F., 2001. Commentary: Is vitamin A playing a hidden role in children's lung function?. Int. J. Epidemiol., 30: 465-466.
CrossRefDirect Link - Serghides, L. and K.C. Kain, 2002. Mechanism of protection induced by vitamin A in Falciparum malaria. Lancet, 359: 1404-1406.
CrossRefDirect Link - Singh, V. and K.P. West, Jr., 2004. Vitamin A deficiency and xerophthalmia among school-aged children in Southeastern Asia. Eur. J. Clin. Nutr., 58: 1342-1349.
CrossRefPubMedDirect Link - Smith, F.R., R. Suskind, O. Thanagkul, C. Leitz and O.R. Goodman, 1975. Plasma vitamin A, retinol binding protein and pre-albumin concentrations in protein-calorie malnutrition. Am. J. Clin. Nutr., 28: 732-738.
Direct Link - Solomons, N.W., 1998. Plant sources of vitamin A and human nutrition: Red palm oil does the job. Nutr. Rev., 56: 309-311.
CrossRefDirect Link - Solomons, N.W. and M. Orozco, 2003. Alleviation of vitamin A deficiency with palm fruit and its products. Asia Pac. J. Clin. Nutr., 12: 373-384.
PubMedDirect Link - Sommer, A., J. Kartz and I. Tarwotjo, 1984. Increased risk of respiratory disease and diarrhea in children with preexisting mild vitamin A deficiency. Am. J. Clin. Nutr., 40: 1090-1095.
Direct Link - Spears, K., C. Cheney and J. Zerzan, 2004. Low plasma retinol concentrations increase the risk of developing bronchopulmonary dysplasia (BPD) and long-term respiratory disability very- low-birth-weight infants. Am. J. Clin. Nutr., 80: 1589-1594.
Direct Link - Spinnler, B.A., 2003. A place for palm fruit to eliminate vitamin A deficiency. Asia Pacific J. Clin. Nutr., 12: 369-372.
PubMedDirect Link - Strube, Y.N.J., J.L. Beard and A.C. Ross, 2002. Iron deficiency and marginal vitamin A deficiency affected growth, hematological indices and the regulation of iron metabolism genes in rats. J. Nutr., 132: 3607-3615.
Direct Link - Suharno, D., C.E. West, Muhalil, D. Karyadi and J.G.A.G. Hautvast, 1993. Supplementation with vitamin A and iron for nutritional anaemia in pregnant women in West Java, Indonesia. Lancet, 342: 1325-1328.
Direct Link - Thatcher, A.J., C.M. Lee and J.W. Erdman, 1998. Tissue stores of β-carotene are not conserved for later use as a source of vitamin A during compromised vitamin A status in Mongolian gerbils (Meriones unguiculatus). J. Nutr., 128: 1179-1185.
PubMedDirect Link - Thurnham, D.I., C.A. Northrop-Clewes, F.S.W. McCullough, B.S. Das and P.G. Lunn, 2000. Innate immunity, gut integrity and vitamin A in Gambian and Indian infants. J. Infant Dis., 182: S23-S28.
PubMedDirect Link - Van Poppel, G., 1993. Carotenoids and cancer: An update with emphasis on human intervention studies. Eur. J. Cancer, 29: 1335-1344.
CrossRef - Van Vliet, T., M.F. Van Vlissingen, F. Van Schaik and H. Van den Berg, 1996. Beta-carotene absorption and cleavage in rats is affected by the vitamin A content of the diets. J. Nutr., 126: 499-508.
PubMedDirect Link - Vijayaraghavan, K., U. Nayak, M.S. Bamji, G.N.V. Ramana and V. Reddy, 1997. Home gardening for combating vitamin A deficiency in rural India. Food Nutr. Bull., 18: 102-102.
Direct Link - Vuong, L.T., S.R. Dueker and S.P. Murphy, 2002. Plasma β-carotene and retinol concentrations of children increase after a 30-day supplementation with the fruit Momordica cochinchinensis (gac). Am. J. Clin. Nutr., 75: 872-879.
PubMedDirect Link - Wang, T.W., H. Zhang and J.M. Parent, 2005. Retinoic acid regulates postnatal neurogenesis in the murine subventricular zone-olfactory tube bulb pathway. Development, 132: 2721-2732.
CrossRefDirect Link - West, K.P., 2002. Extent of vitamin A deficiency among preschool children and women of reproduction age. J. Nutr., 132: 2857S-2866S.
PubMedDirect Link - Wiedermann, U., L.A. Hanson, J. Holmgren, H. Kahu and U.I. Dahlgren, 1993. Impaired mucosal antibody response to the cholera toxin in vitamin A deficient rats immunized with oral cholera vaccine. Infect. Immun., 61: 3952-3957.
PubMedDirect Link - Ye, X., S. Al-Babili, A. Klöti, J. Zhang, P. Lucca, P. Beyer and I. Potrykus, 2000. Engineering the provitamin A (β-Carotene) biosynthetic pathway into (Carotenoid-free) rice endosperm. Science, 287: 303-305.
CrossRefDirect Link - Zhao, Z. and A.C. Ross, 1995. Retinoic acid repletion restores the number of leukocytes and their subsets and stimulates natural cytotoxicity in vitamin A-deficient rats. J. Nutr., 125: 2064-2073.
PubMedDirect Link