
Budianti Kadidaa
Department of Agriculture Sciences, Faculty of Postgraduate Program, Halu Oleo University, Kendari, Indonesia
Gusti Ray Sadimantara
Department of Agrotechnology, Faculty of Agriculture, Halu Oleo University, 93232 Kendari, Southeast Sulawesi, Indonesia
LiveDNA: 62.19189
Suaib
Department of Agrotechnology, Faculty of Agriculture, Halu Oleo University, 93232 Kendari, Southeast Sulawesi, Indonesia
La Ode Safuan
Department of Agrotechnology, Faculty of Agriculture, Halu Oleo University, 93232 Kendari, Southeast Sulawesi, Indonesia
Muhidin
Department of Agrotechnology, Faculty of Agriculture, Halu Oleo University, 93232 Kendari, Southeast Sulawesi, Indonesia
LiveDNA: 62.16775
Asian Journal of Crop Science
Year: 2017 | Volume: 9 | Issue: 4 | Page No.: 109-117







ABSTRACT
Background and Objective: Rice is the major cereal crop for the people of North Buton and Southeast Asia, as well. To determine the diversity of various local upland rice genotypes based on agronomic traits and yield potential, a field experiment was conducted in villages of the Kambowa district, North Buton Regency, Indonesia. Materials and Methods: Twenty-two local upland rice genotypes were evaluated for their diversity in regard to agronomic traits and yield potential. The experimental plots were laid out in a randomized complete block design (RCBD) with 3 replicates. Statistical analysis of variance (two-way ANOVA) to test the significance and to measure the least significant differences between means was carried out. Results: Based on plant height, 12 cultivars were classified as tall plants and 10 cultivars were classified as short plants. The tallest upland rice plants were Wakawondu and Wangkaluku, while the Wajini cultivar was the shortest one. Among the 22 local upland rice cultivars, 7 genotypes were recorded as having a potential yield higher than 3.0 t ha–1. The grain yield/plant exhibited positive significant correlations with filled grain number (r = 0.910), 1000-grain weight (r = 0.906), grain number (r = 0.890), tiller number (r = 0.753), productive tiller number (r = 0.747) and panicle length (r = 0.635) at a 0.01 significance level. However, plant height and leaf area had a negative correlation with the yield/plant (r = -0.140 and r = -0.092, respectively). Furthermore, yield was observed to be positively associated with panicle bearing tillers and number of filled grains/panicle and these characteristics were noticed to exert strong direct effects on grain yield/plant. Conclusion: It was concluded that filled grain number, 1000-grain weight, grain number/panicle, productive tiller number and panicle length can be considered good criteria for the selection of rice cultivars suitable for crop yield improvement programs.
PDF Abstract XML References Citation
Received: July 19, 2017;
Accepted: September 25, 2017;
Published: October 05, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Budianti Kadidaa, Gusti Ray Sadimantara, Suaib, La Ode Safuan and Muhidin, 2017. Genetic Diversity of Local Upland Rice (Oryza sativa L.) Genotypes Based on Agronomic Traits and Yield Potential in North Buton, Indonesia. Asian Journal of Crop Science, 9: 109-117.
DOI: 10.3923/ajcs.2017.109.117
URL: https://scialert.net/abstract/?doi=ajcs.2017.109.117
DOI: 10.3923/ajcs.2017.109.117
URL: https://scialert.net/abstract/?doi=ajcs.2017.109.117
INTRODUCTION
Rice (Oryza sativa L.) is the main food for more than 2.5 billion people in the world and it covers 9% of the earth's arable land. The increased demand for rice is expected to enhance production in many parts of Asia, Africa and Latin America1. It supplies 21% of the global human energy/capita and 15% of the per capita supply of protein. Asian countries contribute over 90% of the world's production of rice2. Rice grain contains 75-80% starch, 12% water and 7% protein3,4. The growth of upland rice is affected by genetic factors and is also strongly influenced by environmental factors in which it grows.
Rice is very important as a main staple food. Indonesia is included as a major rice producer and as the main consumer5,6. The demand for rice is expected to increase in the future and rice production must increase in line with population growth. The Indonesian government has two policies to reach rice self-sufficiency, through the increase of rice production and the reduction of rice consumption, while promoting other source staple foods such as sago7, cassava or corn6. Rice production should be increased through the extension of new paddy fields and increased productivity on existing wetlands5,6.
One effort to comply with rice production policies is to utilize dry land that is still wide enough and to focus on its development. Dry land in Indonesia is estimated to occupy approximately 60.7 million ha or 88.6% of the land area, spread throughout wet climates dominated by Ultisol, which has fewer nutrients and high levels of acidity. Additionally, the content of organic matter easily decreases if the soil is used8, nevertheless, there is general potential for the development of upland rice in these areas. The upland marginal potential for the development of food crops reached an area of 35 million out of 50 million ha9.
Increased upland rice productivity through plant breeding programs aims to obtain a high yield of superior cultivars. One of the stages before a cultivar is detachable is multi-location trials. From the multi-location tests, results are expected to be obtained for cultivars that are adapted to specific environments and have good stability in specific environments or different environments, so they can be released as new cultivars. Adaptability and stability of yield performance are important factors to consider in choosing the varieties to recommend to the farmers10.
The formation of superior cultivars requires the consideration of the stability of the results in a systematic and continuous nature, ranging from the establishment of the base population to testing11. The results reveal important criteria for evaluating the adaptability and yield stability of a cultivar. Measurement of the relative stability of a cultivar on a wide range of areas is important to determine the efficiency of testing in various environments, as in Indonesian upland rice growing environments, which include a wide variety of types of land use, soil types, methods of cultivation, cropping patterns and growing seasons.
Until now, the superior cultivar has been a greater contributor to the increase in productivity than other technological components, although each cultivar has different capabilities in different environmental conditions. This causes the cultivars to have differences in yield if planted in different environments4,12. Ecological factors can alter the yield potential of upland rice cultivars13. The development of superior cultivars needs to be assessed through the interaction of cultivars and the environment to determine the yield potential of a cultivar in different environments. The appearance of plants is affected by factors such as the cultivar, the environment and the interaction between the cultivar and the environment. All three of these factors contribute to variable results with genotype x environment interactions14,15. Cultivar and environment interactions are important for the study of plant breeders in developing cultivars for selection because cultivars will react differently to different environments. Furthermore, sustain ability of rice production depends on the development of new rice cultivars with high yield and stable performance across different environments16. Various types of upland rice have been cultivated by the people in the North Buton Regency. In addition to the different types of rice that are currently being cultivated, there is the possibility to cultivate other types that have superior agronomic characteristics. The objective of the present study was to characterize 22 upland rice cultivars based on agronomic traits and yield potential to provide useful information to facilitate the choice of genitors for rice plant breeding programs to develop new high-yield varieties.
MATERIALS AND METHODS
The experiment was conducted on marginal land at 2 locations (Kambowa and Eelahaji villages) in North Buton. Kambowa is located between 5°01’34.461S and 122°56’59.668E at an altitude of 26.18 m above sea level, while Eelahaji is located between 4o41’15.737S and 123°07’54.124E at an altitude of 36.27 m above sea level. The research was carried out in the rainy season. The materials used in this study were seeds of local upland rice cultivars of North Buton obtained from preliminary studies and organic manure that was applied during the final land preparation.
The experimental plots were laid out in a randomized complete block design (RCBD). The field was divided into three blocks, representing 3 replicates. Twenty-two cultivars were distributed to each plot within each block randomly. The seeds were directly grown in the soil plots. The local upland rice cultivars consisted of Wakombe (V1), Wabalongka (V2), Warumbia Merah (V3), Patirangga (V4), Wa Apolo (V5), Kasakabari (V6), Wangkaluku (V7), Wangkariri (V8), Warangka (V9), Wabila Kambawa (V10), Wakawondu (V11), Wajini (V12), Wawonii (V13), Mantebeka (V14), Wampogeru (V15), Wabila Lambale (V16), Warumbia Putih (V17), Watanta (V18), Waburiburi (V19), Warara (V20), Wankatema (V21) and Warema (V22). Land preparation for the planting of seeds involved slashing of the bush, plowing and harrowing. The seeds were planted at a rate of three seeds/hole at a depth of 5 cm, with an inter and intra-row spacing of 25×25 cm. The plot size was 3×4 m2.
Several agronomic traits, i.e., plant height, leaf width, leaf length, flag leaf length, tiller number and productive tiller number were observed to characterize the cultivars phenotypically. Data on grain yield and some other yield components, i.e., harvesting date, number of grains/panicle, filled grains and unfilled grains, 1000-grain weight and total grain yield (t ha–1) at 14% moisture level were collected.
Statistical analyses: Statistical analyses were performed using two-ways of analysis of variances (ANOVA) by means using the Statistical Package of Social Sciences (SPSS) program version 20 for Windows (Chicago, IL, USA). If the test result showed a significant difference, then tests of treatment differences were performed using Duncan’s multiple range test (DMRT) and Pearson’s correlation coefficient analysis. Differences of p<0.05 were considered statistically significant.
RESULTS AND DISCUSSION
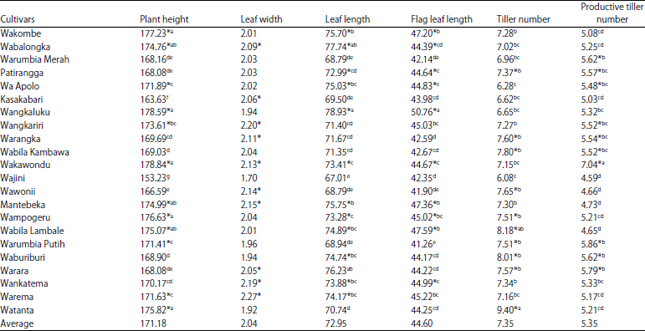
Characteristics of various rice growth components: Based on the observations of various growth components of local upland rice cultivars of North Buton, the average plant height ranged from 153.23-178.84 cm (Table 1). The large difference in the plant height averages of some cultivars tested was due to genetic differences. The differences in plant height were determined more by genetic factors. In addition to being influenced by genetic factors, plant height was also influenced by environmental growth conditions17. Furthermore, Bhadru et al.18, stated that plant height was highly correlated with the level of lodging and ease of harvest, so it was one of the important characteristics that influences the levels of acceptance of new cultivars by farmers. Generally, farmers were less fond of strong plant height performance of cultivars due to vulnerable falls, while too-short cultivars (less than 80 cm) were often difficult to harvest.
In these observations, 12 cultivars were classified as tall plants and 10 cultivars were classified as short plants (Table 1). The tallest local upland rice cultivars were Wakawondu and Wangkaluku, while the Wajini cultivar was the shortest rice plant. According to Baihaki and Wicaksana19 the size of the effect of genotype×environment interactions highly depended on cultivar variability and complexity of the environment that influences it. If the environmental growth suited the plant growth, it could increase plant height and crop production.
The leaves are part of plant organs that are important for the ongoing process of photosynthesis, so the greater the leaf surface area, the higher the products of photosynthesis produced by the plant. However, if the number of leaves was too high and the leaves shaded each other, there would be a significant negative effect on the plant because the shaded leaves would use the photosynthate that was produced by leaf shade.
The results of some local rice cultivars of North Buton (Table 1) showed that there were 10 cultivars that had leaves wider than the average value of rice leaf area. Plants that had the widest leaves were the Warema and Wankatema cultivars, while cultivars that had more narrow leaves were the Wajini, Waburiburi and Wangkaluku cultivars but these cultivars produced more tillers and productive tillers. The tiller numbers of all the local upland rice cultivars tested ranged from 6.08-9.40 with an average value of 7.35 tillers.
The fewest tillers (6.08) of upland rice were produced by the Wajini cultivar. This cultivar also produced few productive tillers (4.59) and the most were produced by the Wakawondu cultivar that produced as many as 7.04 productive tillers. According to Wang et al.20, variation in grain development and yield among tillers was variable over the varieties that differed in tillering ability. Meanwhile, variation in grain yield and quality among tillers has been considered as a major factor affecting yield potential and quality for a given rice variety. Gunasekaran et al.21, stated that productive tillers, followed by filled grains/panicle and 1000-grain weight, were important characteristics for improving the yield potential of rice.
Top of form: The flag leaf is a plant part that was important to observe because the position of the flag leaf is closest to the panicle of the rice plant, so it becomes the most important source of photosynthate for grain growth and development in rice plants.
| Table 1: | Rice growth components of 22 local upland rice cultivars of North Buton |
 | |
Averages of plant height (cm), leaf width (cm), leaf length (cm), flag leaf length (cm), tiller number and productive tiller number. The numbers followed by an asterisk (*) show a greater value than the average value. Means followed by the same letter are not significantly different at the 5% level by DMRT | |
The result of leaf length and flag leaf length showed that longer leaves (78.93 cm) and flag leaves (50.76 cm) were produced by the Wangkaluku cultivar. In contrast, this cultivar produced lower than average numbers of tillers and productive tillers (Table 1).
The results of the evaluation of all growth parameters, as shown in Table 1, showed that the Wangkaluku cultivar produced values consistently greater than the average value for each growth variable, except for the number of tillers (7.15). Based on these data, it can be argued that the cultivars that have high performance and long leaves will produce fewer numbers of tillers and productive tillers. It might be that most of the photosynthate produced are widely used for the growth of stems and leaves of the plant, so the plant does not produce more tillers and productive tillers. Recently, Wang et al.22, reported that the unequal distribution of photosynthetically active radiation (PAR) was the source of heterogeneity in an individual’s tiller yields, in which early emerging superior tillers preempted the uppermost light source and shaded the late emerging tillers under limited light conditions.
The number of productive tillers tested in the present study ranged from 4.59-7.04 productive tillers with an average value of 5.35 tillers. There were 10 cultivars with productive tiller numbers above the average, i.e., Warumbia Merah, Patirangga, Wa Apolo, Wangkariri, Warangka, Wabila Kambawa, Wakawondu, Warumbia Putih, Waburiburi and Warara cultivars. The difference in productive tiller numbers may be affected by environmental factors. According to Anandan et al.23, that the environmental conditions vary from one place to another and that the crop needs specific environmental conditions will lead to a diversity in plant growth.
Characteristics of various rice yield components: Observations of the yield components included the time to harvest (days), the number of grains (grains/panicle), the number of filled grains (grain/panicle), the number of unfilled grains (grains/panicle), the 1000-grain weight (g) and the grain production (t ha–1). Harvesting was set according to the number of days after planting. Harvesting time ranged between 131-143 days.
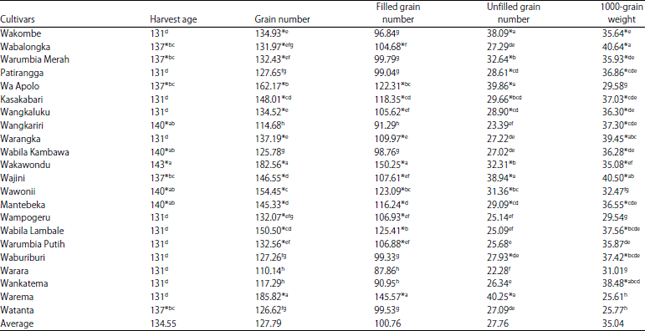
The results indicated that the local upland rice cultivar of North Buton with the longest harvesting time was Wakawondu, with a harvesting time of 143 days after planting. Furthermore, the cultivars with 140 days of harvesting time were the Wangkariri, Wabila Kambowa, Wawonii and Mantebeka cultivars. The results of the various yield components are shown in Table 2. According to Ranawake et al.15, rice yield has a positive correlation with grain-filling period after flowering because the plants should accumulate sufficient vegetative growth to support their reproductive phase.
| Table 2: | Rice yield components of 22 local upland rice cultivars of North Buton |
 | |
Averages of harvest age (day), grain number (grain/panicle), filled grain number (grain/panicle), unfilled grain number (grain/panicle) and 1000-grain weight (g), the numbers followed by an asterisk (*) showed a greater value than the average value, Means followed by the same letter are not significantly different at the 5% level by DMRT | |
On the other hand, Cho et al.24, stated that the duration of seed filling generally ranges from 5-20 days, followed by a maturation phase of approximately 20 days. Estimated harvesting time ranges from 25-40 days after flowering.
The results of the grain numbers of various local North Buton upland rice cultivars grown in Kambowa village (Table 2), showed that the grain number ranged from 110.14-185.82 grains/panicle with an average value of 127.79 grains/panicle. Most of the local upland rice cultivars produced filled grains higher than the average value, except 7 cultivars, i.e., Patirangga (127.65), Waburiburi (127.26), Watanta (126.62), Wabila Kambawa (125.78), Wankatema (117.29), Wangkariri (114.68) and Warara (110.14), which produced grain numbers/panicle lower than the average. Three major components, panicle number/plant, grain number/panicle and grain weight, determined rice yield production. Grain weight is associated with grain size and shape, which were defined as grain length, grain width and grain thickness25.
The results of the filled grain numbers showed that there were 8 cultivars that produced filled grains higher than the average value of filled grain in the cultivars tested. The average yield of filled grain ranged from 87.86-150.25 grains/panicle. The upland rice cultivar that produced the highest number of filled grains was Wakawondu (150.25 grains/panicle) and the lowest (87.86 grains/panicle) was the Warara cultivar. Observational results recorded that Wabalongka, Kasakabari, Wa Apolo, Wangkaluku, Warangka, Wakawondu, Mantebeka, Wabila Lambale and Warema produced more filled grain content compared with the average value. The percentage of filled grains indicates panicle fertility rates that are calculated based on the total number of grains/panicle. The number of filled grains support the yield potential of rice, so the higher panicle fertility, indicated by the percentage of filled grains, will be more likely to provide high yields26.
The percentage of unfilled grains was also one agronomic characteristic that affected rice yield. The emptiness of grains can be caused by plant genetic factors and by environmental factors such as drought, excess or lack of nutrients, pest and disease attacks that occur during the seed filling process. An increase in the amount of unfilled grain produced will cause a decrease in yield production of a rice cultivar.
In the present study, the unfilled grain numbers ranged from 22.28-40.25 grains/panicle, with an average value of 27.76 grains/panicle (Table 2). The Warara cultivar produced the lowest number of unfilled grains (22.28 grains/panicle), while the Warema cultivar produced the highest number of unfilled grains (40.25 grains/panicle).
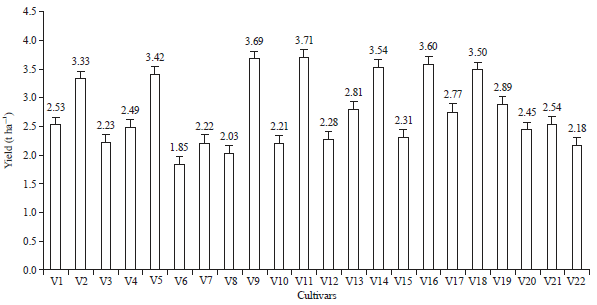 | |
| Fig. 1: | Yield potential (t ha–1) of 22 local upland rice genotypes |
| Bars represent SE | |
This shows that the growth and production of plants and plant age were more affected by genetic factors than the plant’s growth environment.
The cultivars that produced unfilled grain numbers higher than the average value consisted of Wakombe, Wa Apolo, Warumbia Merah, Wakawondu, Wawonii, Wajini and Warema. One of the agronomic characteristics that will affect rice yield is the percentage of unfilled grain. High or low numbers of unfilled grains/panicle, besides being caused by genetic factors, were also caused by environmental factors including drought, excess or lack of nutrients, pest and disease attacks during the process of fertilization and seed filling. An increase in the number of unfilled grains produced will cause a decrease in rice yield produced by a cultivar.
The observation of various upland rice cultivars (Table 2) showed that the average weight of 1000 grains was 35.04 g with a range from 25.61-40.64 g. The weight of 1000 grains indicates the size of the seeds produced by rice plants. The 1000-grain weight was one of the most important yield components after the number of filled grains, spikelet number, density, seed setting rate and panicle length27. According to Kato28, Li et al.29 and Kato and Horibata30, the 1000-grain weight was positively correlated with the size of the grain, i.e., length, width and seed filling number. These results indicated that the upland rice cultivars have a high fertility rate.
Seven genotypes among 22 local upland rice genotypes were recorded as having a yield potential (t ha–1) higher than 3.0 t ha–1 (Fig. 1). The genotypes consisted of Wakawondu, Warangka, Wabila Lambale, Mantebeka, Watanta, Wa Apolo and Wabalongka. The lowest yield was produced by Kasakabari. Rice yield was a complex trait that was directly associated with the grain size, the panicle number and the number of grains/panicle31. Rice yield has been determined not only by spikelet number, grain weight and number of grains/panicle but also by plant height and flowering time32. Furthermore, yield, plant height and flowering time were the three most important agronomic traits in rice33. Days to flowering affects the plant height and the yield of the rice plant and was the most important factor of the varietal selection criteria.
Pearson’s correlation coefficients among yield and yield components of local upland rice cultivars were calculated (Table 3) using SPSS software (version 20.0). The grain yield/plant exhibited positive significant correlations with filled grain number (r = 0.910), 1000-grain weight (r = 0.906), grain number (r = 0.890), tiller number (r = 0.753), productive tiller number (r = 0.747) and panicle length (r = 0.635) at the 0.01 significance level (Table 3). Positive and significant correlations of panicle length, flag leaf area and number of grains/panicle have been previously reported17. Furthermore, grain yield was significantly correlated with number of tillers per plant, number of filled grains/panicle and 1000-grain weight34,35. In the present study, plant height and leaf area were reported to have a negative correlation with yield/plant (r = -0.140 and r = -0.092, respectively) at the 0.01 significance level. Hairmansis et al.26, also observed that final grain yield was negatively correlated with plant height. Filled grain number was positively and significantly correlated with yield/plant (α = 0.01, r = 0.910). This finding was aligned with the findings of Luzikihupi36 and Bhatti et al.37. Takai et al.38 and Zhao et al.39, which stated that rice grain filling was a critical and dynamic factor that determines the grain yield.
| Table 3: | Pearson’s correlation coefficients among yield and yield component characteristics of evaluated local upland rice cultivars |
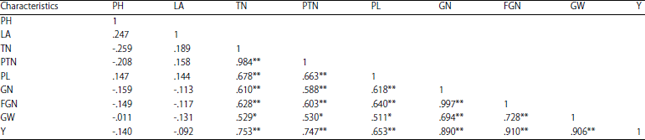 | |
PH: Plant height, LA: Leaf area, TN: Tiller number, PTN: Productive tiller number, PL: Panicle length, GN: Grain number, FGN: Filled grain number, GW: 1000-grain weight, Y: Yield/plant. **Correlation is significant at the 0.01 level (2-tailed), *Correlation is significant at the 0.05 level (2-tailed) | |
Many factors affect filled grain percentage, such as climate, soil, variety, fertilizer application and insect and pest attacks40. The 1000-grain weight was positively and significantly correlated with the yield/plant (α = 0.01, r = 0.906). Previously, Prasad et al.41 and Bhatti et al.37, also reported the correlation between the 1000-grain weight and the yield/plant. Further, Ashraf et al.42, reported that the 1000-grain weight was a genetic characteristic least influenced by the environment.
CONCLUSION
The result of study showed that 7 rice genotypes from North Buton have better growth characteristic and high yielded potential. The present study highlighted the existence of diversity among the 22 local upland rice cultivars, where seven rice genotypes, i.e., Wakawondu, Warangka, Wabila Lambale, Mantebeka, Watanta, Wa Apolo and Wabalongka, have favorable growth characteristics and high yield potential. These cultivars can be selected for adaptability tests to become a recommended variety. Based on Pearson’s correlation analysis, grain yield is significantly and highly correlated with filled grain number (r = 0.910), 1000-grain weight (r = 0.906), grain number (r = 0.890), tiller number (r = 0.753), productive tiller number (r = 0.747) and panicle length (r = 0.635) at the 0.01 significance level. However, plant height and leaf area were negatively correlated with yield/plant (r = -0.140 and r = -0.092, respectively). Further, yield was observed to be positively associated with panicle bearing tillers and number of filled grains/panicle and these characteristics were observed to exert strong direct effects on grain yield/plant. Filled grain number, 1000-grain weight, grain number/panicle, productive tiller number and panicle length can be considered good criteria for the selection of rice cultivars suitable for crop yield improvement programs.
SIGNIFICANCE STATEMENTS
This study discovered the possible high yield potential of local upland rice that can be beneficial for improving rice yield on dry land. This study help researchers to uncover the critical factors of the potential production of upland rice on marginal land that many researchers were not able to explore previously. The result revealed that the high yield potential and stability of upland rice cultivars may be attained in discovering new promising cultivars with high yield potential and stability in marginal land.
REFERENCES
- Devi, A.S., V. Krishnasamy, M. Raveendran, N. Senthil and S. Manonmani, 2011. Molecular and genetic dissection of peduncle elongation in rice. Electron. J. Plant Breed., 2: 1-7.
Direct Link - Oko, A.O., B.E. Ubi, A.A. Efisue and N. Dambaba, 2012. Comparative analysis of the chemical nutrient composition of selected local and newly introduced rice varieties grown in Ebonyi State of Nigeria. Int. J. Agric. Forest., 2: 16-23.
CrossRefDirect Link - Hossain, S., M. Haque and J. Rahman, 2015. Genetic variability, correlation and path coefficient analysis of morphological traits in some extinct local Aman rice (Oryza sativa L.). J. Rice Res. Vol. 3.
CrossRef - Muhidin, K. Jusoff, S. Elkawakib, Y. Musa and Kaimuddin et al., 2013. The development of upland red rice under shade trees. World Applied Sci. J., 24: 23-30.
Direct Link - Sutariati, G.A.K., A. Khaeruni, Y.B. Pasolon, Muhidin and L. Mudi, 2016. The effect of seed bio-invigoration using indigenous Rhizobacteria to improve viability and vigor of upland rice (Oryza sativa L.) seeds. Int. J. PharmTech Res., 9: 565-573.
Direct Link - Muhidin, S. Leomo, S. Alam and T. Wijayanto, 2016. Comparative studies on different agroecosystem base on soil physicochemical properties to development of sago palm on dryland. Int. J. ChemTech Res., 9: 511-518.
Direct Link - Sondari, N. and E.S. Nurkhalidah, 2012. Application of bokashi botom ash for increasing upland rice yield and decreasing grain Pb content in vitric hapludans. J. Trop. Soils, 17: 157-163.
Direct Link - Sadimantara, G.R., Muhidin and E. Cahyono, 2014. Genetic analysis on some agro-morphological characters of hybrid progenies from cultivated paddy rice and local upland rice. Adv. Stud. Biol., 6: 7-18.
Direct Link - Palanog, A.D., C.A. Endino, I.M.G. Ciocon, L.T. Ines and E.M. Libetario, 2014. Adaptability and stability analysis of newly-released rice varieties using GGE biplot analysis. Asia Life Sci., 23: 515-527.
Direct Link - Venuprasad, R., M.T. Sta Cruz, M. Amante, R. Magbanua, A. Kumar and G.N. Atlin, 2008. Response to two cycles of divergent selection for grain yield under drought stress in four rice breeding populations. Field Crops Res., 107: 232-244.
CrossRefDirect Link - Allard, R.W. and A.D. Bradshaw, 1964. Implications of genotype-environmental interactions in applied plant breeding. Crop Sci., 4: 503-508.
Direct Link - Ranawake, A.L. and U.G.S. Amarasinghe, 2014. Relationship of yield and yield related traits of some traditional rice cultivars in Sri Lanka as described by correlation analysis. J. Sci. Res. Rep., 3: 2395-2403.
Direct Link - Akter, A., J.M. Hassen, U.M. Kulsum, M.R. Islam, K. Hossain and M.M. Rahman, 2014. AMMI biplot analysis for stability of grain yield in hybrid rice (Oryza sativa L.). J. Rice Res., Vol. 2.
CrossRef - Khan, A.S., M. Imran and M. Ashfaq, 2009. Estimation of genetic variability and correlation for grain yield components in rice (Oryza sativa L.). Am. Eurasian J. Agric. Environ. Sci., 6: 585-590.
Direct Link - Bhadru, D., R.D. Bhadru, M.S. Ramesha, 2011. Correlation and path coefficient analysis of yield and yield contributing traits in rice hybrids and their parental lines. Electron. J. Plant Breeding, 2: 112-116.
Direct Link - Wang, F., F.M. Cheng and G.P. Zhang, 2007. Difference in grain yield and quality among tillers in rice genotypes differing in tillering capacity. Rice Sci., 14: 135-140.
CrossRefDirect Link - Gunasekaran, M., N. Nadarajan and S.V.S.R.K. Netaji, 2010. Character association and path analysis in inter-racial hybrids in rice (Oryza Sativa L.). Electron J. Plant Breed., 1: 956-960.
Direct Link - Wang, Y., T. Ren, J. Lu, R. Ming and P. Li et al., 2016. Heterogeneity in rice tillers yield associated with tillers formation and nitrogen fertilizer. Agron. J., 108: 1717-1725.
CrossRefDirect Link - Anandan, A., G. Rajiv, R. Eswaran and M. Prakash, 2011. Genotypic variation and relationships between quality traits and trace elements in traditional and improved rice (Oryza sativa L.) genotypes. J. Food Sci., 76: H122-H130.
CrossRefDirect Link - Cho, D.S., S.K. Jong, S.Y. Son and Y.K. Park, 1988. Studies on the duration and rate of grain filling in rice (Oryza sativa L.) II. Difference between the parts of a panicle. Korean J. Crop Sci., 33: 5-11.
Direct Link - Duan, P., Y. Rao, D. Zeng, Y. Yang and R. Xu et al., 2014. SMALL GRAIN 1, which encodes a mitogen‐activated protein kinase kinase 4, influences grain size in rice. Plant J., 77: 547-557.
CrossRefDirect Link - Hairmansis, A., B. Kustianto, Supartopo and Suwarno, 2010. Correlation analysis of agronomic characters and grain yield of rice for tidal swamp areas. Indonesian J. Agric. Sci., 11: 11-15.
Direct Link - Liu, E., Y. Liu, G. Wu, S. Zeng and T.G.T. Thi et al., 2016. Identification of a candidate gene for panicle length in rice (Oryza sativa L.) via association and linkage analysis. Front. Plant Sci., Vol. 7.
CrossRefDirect Link - Kato, T., 2010. Variation and association of the traits related to grain filling in several extra-heavy panicle type rice under different environments. Plant Prod. Sci., 13: 185-192.
CrossRefDirect Link - Li, Y., C. Fan, Y. Xing, Y. Jiang and L. Luo et al., 2011. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet., 43: 1266-1269.
CrossRefDirect Link - Kato, T. and A. Horibata, 2011. Non-random distribution of the alleles for good grain filling at OsAGPS2 and OsSUT1 among a wide range of rice (Oryza sativa L.) cultivars. Breed. Sci., 61: 217-220.
CrossRefDirect Link - Xing, Y. and Q. Zhang, 2010. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol., 61: 421-442.
CrossRefPubMedDirect Link - Wei, X., J. Xu, H. Guo, L. Jiang and S. Chen et al., 2010. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol., 153: 1747-1758.
PubMedDirect Link - Xue, W.Y., Y. Xing, X. Weng, Y. Zhao and W. Tang et al., 2008. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet., 40: 761-767.
CrossRefDirect Link - Sreedhar, S., T.D. Reddy and M.S. Ramesha, 2011. Genotype x environment interaction and stability for yield and its components in hybrid rice cultivars (Oryza sativa L.). Int. J. Plant Breeding Genet., 5: 194-208.
Direct Link - Osman, K.A., A.M. Mustafa, F. Ali, Z. Yonglain and Q. Fazhan, 2012. Genetic variability for yield and related attributes of upland rice genotypes in semi arid zone. Afr. J. Agric. Res., 7: 4613-4619.
Direct Link - Luzikihupi, A., 1998. Interrelationship between yield and some selected agronomic characters in rice. Afr. Crop Sci. J., 6: 323-328.
Direct Link - Takai, T., Y. Fukuta, T. Shiraiwa and T. Horie, 2005. Time-related mapping of quantitative trait loci controlling grain-filling in rice (Oryza sativa L.). J. Exp. Bot., 56: 2107-2118.
CrossRefDirect Link - Zhao, B.H., P. Wang, H.X. Zhang, Q.S. Zhu and J.C. Yang, 2006. Source-sink and grain filling characteristics of two-line hybrid rice Yangliangyou 6. Rice Sci., 13: 34-42.
Direct Link - Yoshida, S., 1972. Physiological aspects of grain yield. Ann. Rev. Plant Physiol., 23: 437-464.
CrossRefDirect Link - Ashraf, M., A. Khalid and K. Ali, 1999. Effect of seedling age and density on growth and yield of rice in saline soil. Pak. J. Biol. Sci., 2: 860-862.
Direct Link