
Hassan A. Elfadaly
Department of Zoonotic Diseases, National Research Centre, Dokki, Giza, Egypt
Nawal A. Hassanain
Department of Zoonotic Diseases, National Research Centre, Dokki, Giza, Egypt
Raafat M. Shaapan
Department of Zoonotic Diseases, National Research Centre, Dokki, Giza, Egypt
Mohey A. Hassanain
Department of Zoonotic Diseases, National Research Centre, Dokki, Giza, Egypt
LiveDNA: 20.10587
Ashraf M. Barakat
Department of Zoonotic Diseases, National Research Centre, Dokki, Giza, Egypt
Khaled A. Abdelrahman
Department of Parasitology and Animal Diseases, National Research Centre, Giza, Egypt
LiveDNA: 20.10947
Asian Journal of Epidemiology
Year: 2017 | Volume: 10 | Issue: 1 | Page No.: 37-44







ABSTRACT
Background and objective: Toxoplasma gondii is a zoonotic opportunistic cyst-forming protozoan has usual clonal population of three virulent human and animal genotypes which differ according to their virulence in mice and epidemiological patterns. Animals are of zoonotic impact and regarded as high prevalent sources for human toxoplasmosis via their meat containing viable T. gondii tissue cysts. However, serological assays are not enough for approving virulent T. gondii tissue cysts or confirm virulent types harboring animals so the current study was aimed to evaluate the zoonotic hazard of T. gondii seropositive Egyptian animals and aborted women via mice viability test and genotyping as excellent bio-indicators for virulent strains. Methodology: Both blood and the matching tissue samples were collected from 76 aborted women and 755 farm animals (254 sheep, 293 goats, 45 cows, 41 buffalos, 34 camels and 88 free range poultry). All samples were assayed serologically by using ELISA. The tissue samples which were identical to seropositive sera were digested and microscopically examined besides exposed to DNA detection and genotyping. The microscopic confirmed bradyzoites containing ones were bio-assayed through intra-peritoneal passage in mice as viability test. Results: The total seropositive percentages and the suspected microscopic examined samples plus to DNA percentages detected and succeeded isolates via mice viability test were (6, 8, 4, 0, 0, 2 and 2) corresponding to (women, sheep, goat, cows, buffaloes, camels and free range poultry) respectively. Genotyping confirmed 22 isolates (2 type- I, 13 type- II and 7 type- III). Conclusion: The mice viability and DNA approve along with genotyping were signify T. gondii virulent types 1-3 viable cysts from Egyptian women, animals and poultry and reflect zoonotic bio-hazard of public health concern.
PDF Abstract XML References Citation
Received: September 18, 2016;
Accepted: November 11, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hassan A. Elfadaly, Nawal A. Hassanain, Raafat M. Shaapan, Mohey A. Hassanain, Ashraf M. Barakat and Khaled A. Abdelrahman, 2017. Molecular Detection and Genotyping of Toxoplasma gondii from Egyptian Isolates. Asian Journal of Epidemiology, 10: 37-44.
DOI: 10.3923/aje.2017.37.44
URL: https://scialert.net/abstract/?doi=aje.2017.37.44
DOI: 10.3923/aje.2017.37.44
URL: https://scialert.net/abstract/?doi=aje.2017.37.44
INTRODUCTION
Toxoplasma gondii is an obligate intracellular tissue cyst-forming protozoan, capable of infecting and multiplying in all warm-blooded animals, birds and humans with worldwide distribution1. It exists in three infective stages, the acute tachyzoite stage which is responsible for the materno-fetal pass, the dormant life-persistent tissue cyst containing bradyzoites which forms particularly in neural and muscular tissue including uterus and placenta, while the environment oocysts are developed only in the feline's gut and disseminated in their feces2. Human toxoplasmosis is a typical meat borne zoonosis1, postnatal humans infection is mostly via eating insufficient cooked meat harboring tissue cysts, which could persist viable in animal tissues3. Also, persons or animals can harbor bradyzoites through food or water contaminated with environment oocysts shed in cat feces.
Toxoplasmosis exceeds opportunistic human impact sequence to cerebral4, congenital5 and ocular toxoplasmosis6,7. Also, maternal-fetal pass in pregnant women possible develop abortion, stillbirths, mental retardation, plus neurological sequel of congenital malformations and ocular disorders8, this usually set consequence to estrogen/progesterone shift9 or related to impermanent gravidity hyperglycemia10. Also, may be potential to anti-inflammatory or corticosteroids therapy11. The disease ranks among the 10 most opportunistic pathogens and the forth deadly pathogen in immune-suppressed patients2.
The animals and humans isolates are types, I, II and III which differ according to their virulence in mice and epidemiological patterns12. Type I strain is highly virulent in mice and mainly recorded in patients with ocular toxoplasmosis and more likely to cause severe human toxoplasmosis13,14. Type II strain is of moderate virulence for mice but the most common strain associated with human infections which creates chronic persistent tissue cysts15. Moreover, latent cystogenic (cyst-forming) types I and II strains have been recorded in patients with congenital disease and AIDS patients16,17. Type III has low-virulence for mice and is uncommon human type but the most frequent strain from animals18.
A variety of different methods has been successively used for genotyping of T. gondii isolates with distinct advantages and disadvantages. Early studies of strain typing were based on multilocus enzyme electrophoresis19 and later focused on microsatellite markers14. Also, randomly amplified polymorphic DNA and Restriction Fragment Length Polymorphism (RFLP) analysis of specific genetic loci has been widely used for T. gondii genotyping12.
Unfortunately, inadequate genotyping data of T. gondii isolates in Africa and middle East including Egypt is due to difficulty in obtaining viable isolates because of poor laboratory facilities in most African countries and the information is mostly based on a few PCR-RFLP markers20. Furthermore, there have been insufficient researches on Egyptian meat producing animals and poultry concerning toxoplasmosis21,22. Serological assays aren’t enough for confirming zoonotic biohazards of T. gondii virulent types in animals. Therefore, the present study aimed to evaluate the zoonotic bio-hazard of T. gondii seropositive Egyptian animals and aborted women via mice viability test and genotyping, as excellent bio-indicators for virulent strains having zoonotic impact.
MATERIALS AND METHODS
Blood and tissue samples: Both blood and the matching tissue samples were collected from 76 aborted women, 254 sheep, 293 goats, 45 cows, 41 buffaloes, 34 camels and 88 free range poultry respectively from Cairo, Giza and Qaliubiya governorates (Egypt).
Serological assay: Sera were separated, labeled and kept at -20°C until examined serologically against toxoplasma infection by ELISA which was carried out according to the method described by Aubert et al.23 using soluble crude antigen prepared from Toxoplasma RH strain tachyzoites.
Digestion of meat samples: Tissue samples were prepared as described by Shaapan and Elfadaly24, 20 g of the equivalent tissue samples from diaphragm and thigh muscles collected from animals during slaughtering in abattoirs and collected from various batches of free range poultries from diverse villages in rural areas in Egypt. The tissues were cut into small cubes about 5×5×5 cm and classified into two groups, the 1st was frozen at -80°C for further DNA extraction while the 2nd group was exposed to pepsin digestion and then bio-assayed for both microscopic examination and mice viability test.
Mice viability test: The test procedures were done according to Elfadaly et al.9, a total number of 48 seronegative Swiss Webster Albino mice obtained from Laboratory Animal House, National Research Centre, Egypt. The inoculated mice were followed up daily for any apparent clinical signs of febrile response or acute toxoplasmosis with the exclusion of dead mice before 48 h of inoculation. The predicted signs may be varied between ascites, roughcast hairs, tottering gait, hunched appearance, with evidence of early emaciation and dehydration or death usually within 72-84 h (with highly virulent types) or nervous manifestations and partial off food (with moderately virulent types). Depending on the virulence of the isolate, the procedure will be continued. If ascites occurred, peritoneal exudates were collected from ascetic mice within 72-84 h DPI and microscopic examination for tachyzoites was done. If mice didn’t show ascites they were sacrificed within 15th day post-inoculation by cervical dislocation, according to Ajzenberg et al.14.
Detection of T. gondii by PCR: Part of frozen tissue samples at -80°C without preservatives will be processed for molecular studies and genotyping. Genomic DNA of T. gondii from the collected tissue samples was extracted by a Kit (ViVantis Co., Malaysia) according to the manufacturer instructions. The B1 gene was amplified25, using primers 1 (5ˊ-TCG GAG AGA GAA GTT CGTCGC AT-3ˊ) and 2 (5ˊ-AGC CTC TCT CTT CAA GCA GCG TA-3ˊ). The following reaction mixture was added in a 0.2 mL PCR tubes: DNA template (100 ng μL–1), 10 μL; Taq polymerase (5 u μL–1) 1 μL; 10x enzyme buffer, 2 μL; dNTPs, 0.8 μL; each primer, 1 μL and Bidest. water to 20 μL. The mixture was briefly spine and placed in the thermal cycler (T gradient, Biometra, Germany), which was programmed as follow: Initial denaturing (95°C/2 min) and 40 cycles consisting of denaturing (95°C/1 min), annealing (55°C/30 sec), extension (72°C/45 sec) and final extension (72°C/10 min). The PCR product was electrophoresed at 80 v/15 min and finally examined using UV trans-illuminator. About 100 bp DNA ladder (Finnzymes) was used as a marker. Positive control was DNA from RH strain and negative control was distilled water in presence of primers.
Genetic characterization: Toxoplasma gondii DNA was extracted from tissues as described by Lehmann et al.26. The PCR-RFLP genotypes of the SAG2 locus were used to determine the genetic type. Samples were analyzed at the SAG2 locus by using a nested PCR approach that separately amplified the 5' and 3' ends of the locus. The 5' end of the locus was amplified by standard PCR for 40 cycles with the primers SAG2.F4 (5'-GCTACCTCGAACAGGAACAC-3') and SAG2.R4 (5'-GCATCAACAGTCTTCGTTGC-3') at an annealing temperature of 65°C. The resulting amplification products were diluted 1/10 in water and a second amplification of 40 cycles was performed with the internal primers SAG2.F (5'-GAAATGTTTCAGGTTGCTGC-3') and SAG2.R2 (5'-GCAAGAGCGAACTTGAACAC-3') by using 1 mL of the diluted product as the template. The amplified fragments were purified with Quia Clean (Qiagen Inc.) and digested with Sau3AI. The 3' end of the locus was similarly analyzed with the primers SAG2.F3 (5'-TCTGTTCTCCGAAGTGACTCC-3') and SAG2.R3 (5'-TCAAAGCGTGCATTATCGC-3') for the initial amplifications and the internal primers SAG2.F2 (5'-ATTCTCATGCCTCCGCTTC-3') and SAG2.R (5'-AACGTTTCACGAAGGCACAC-3') for the second round of amplification at an annealing temperature of 63°C. The resulting amplification products were purified with Quia Clean and digested with HhaI. The PCR products and the restriction fragments were analyzed by 2% agarose gel electrophoresis. Restriction digestion of 5'-end-amplified products with Sau3AI distinguished the type III strain from types I and II strains and HhaI digestion of the 3'-end-amplified fragments differentiated types I and III strains from type II strains. In order to avoid possible contamination, several measures, such as separate space to set up PCRs, filter tips, etc., were taken, as well a negative control (no DNA) and positive controls from different strains of T. gondii were used in order to locate any possible contamination.
Ethical approval: The study was approved ethically by the Medical Research Ethics Committee-National Research Centre, Egypt under registration number 1-2/0-2-1.2012.
RESULTS
Table 1 shows that total seropositive percentages were (42.1, 64.2, 43.3, 35.5, 17.1, 26.5 and 37.5), total suspected microscopic examinations were (42.3, 46, 38.6, 12.5, 0, 44.4 and 27.3), DNA detection percentages (32.7, 52.7, 41.7, 18.8, 0, 55.5 and 36.4) and total succeeded isolates via mice viability test (6, 8, 4, 0, 0, 2 and 2) corresponding to (women, sheep, goat, cows, buffaloes, camels and free range poultry) respectively.
Genotyping: The target regions for amplification were shortened to encompass only the relevant polymorphisms. Primers were selected to separately amplify the 5' and 3' ends of the T. gondii SAG2 locus as 241and 221 bp products, respectively. Digestion of the 5' amplification products with Sau3AI distinguished allele 3 (type III strains) from alleles 1 and 2 (type I and II strains) and digestion of the 3' amplification products with HhaI distinguished allele 2 (type II strains) from alleles 1 and 3 (type I and III strains).
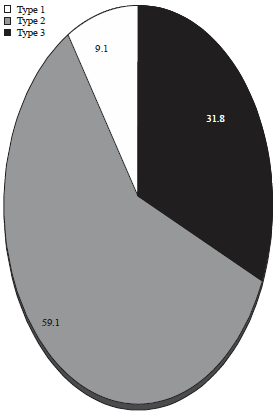 | |
| Fig. 1: | Percentages of genotyped positive samples |
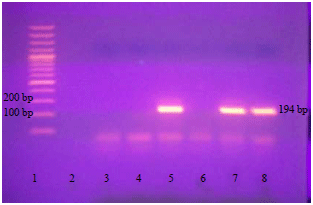 | |
| Fig. 2: | Detection of Toxoplasma genome by PCR using Toxoplasma gene B1 primers |
Lane 1: 100 bp DNA ladder, lanes 7 and 8: Positive isolates (194 bp), lane 5: Reference RH strain, lanes 2, 3, 4 and 6: Negative specimens | |
In this study, samples were typed as 2 (9.1%) type I, 13 (59.1%) type II and 7 (31.8%) type III using PCR-RFLP analysis, the genotypes (Table 1, Fig. 1-4).
Significance and interpretation of the results: This study point out to the zoonotic hazard of T. gondii seropositive Egyptian animals and aborted women via mice viability test and genotyping, as excellent bio-indicators for virulent strains.
| Table 1: | Results of ELISA, microscopic examination, DNA detection, mice viability test and genotyping positive samples |
 | |
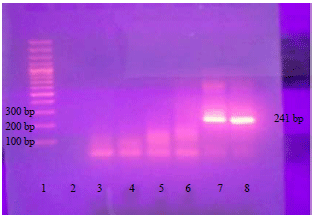 | |
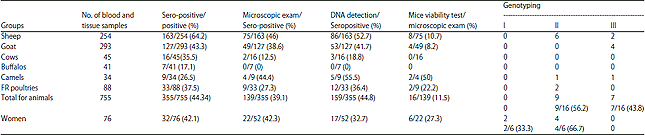
| Fig. 3: | Amplicon of the 5' of the T. gondii SAG2 locus as 241 bp product |
| Lane 1: 100 bp DNA ladder (Fermentas), lanes 7 and 8: Positive isolates (~241 bp), lane 2-6: Negative specimens | |
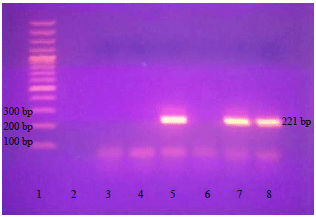 | |
| Fig. 4: | Amplicon of the 3' end of the T. gondii SAG2 locus as 221 bp product |
| Lane 1: 100 bp DNA ladder (Fermentas), lanes 5, 7 and 8: Positive isolates (~221 bp), lanes 2-4 and 6: Negative specimens | |
Bio-assay through intra-peritoneal passage in mice as viability test suspected for T. gondii isolates and DNA percentages were detected. The obtained results concluded that mice viability and DNA approve along with genotyping were signify T. gondii virulent types I, II and III viable cysts from Egyptian women, animals and poultry and reflect zoonotic bio-hazard of public health concern.
DISCUSSION
High prevalence (about 70%) of Egyptians is T. gondii seropositive, perhaps connected to consuming undercooked meat or poultry rather than oocysts contaminated food or water10. So, recent etiological investigation concerning risk factors plus genotyping of T. gondii local Egyptian isolates should be updated to explain the illogic data of high human sero-prevalence. In the present study, B1 gene choice as target to PCR amplification was established on the explanations made by other studies27.
Isolates of T. gondii type-I are poor forming either meat tissue cyst or environmental oocysts (cat feces) and mainly transmitted via placental diffusion, it is highly lethal to mice with high zoonotic concern and likely to cause severe congenital and ocular human toxoplasmosis13. In contrast, latent types II and III are highly cystogenic (cyst-forming) and well shed oocysts. Thus, they essentially spread through meat cysts or sporulated oocysts. Type III is animal strains, rarely seen in human toxoplasmosis28. In the present study, genotyping of 6 T. gondii women isolates confirmed two of type-I (33.3%) and four of type-II (66.7%). Where, 2 isolates of type-I was recorded only in women, denote great prospect placental transmission. A study in 2008 at Ain Shams University Hospitals on 38 female patients with abortion and intrauterine fetal death, genotyping proved 38 isolates, type II was the most prevalent genotype found in 33 (87%). Type I was found in 5 (13%) of the isolates, whereas genotype III was not never found29.
In the existing study, higher possibility of zoonotic transmission was established. Where, genotype-II which is the main women isolates has been detected in meat of Egyptian producing animals (6/8 in sheep, 1/2 in camels and 2/2 in poultry). Thus, mutton in the current study represents considerable zoonotic risk. While free range poultry indicated the probable higher zoonotic risk through 2 isolates of genotype-II. The results of this study agree with Dubey et al.21 who detected high prevalence in Egyptian free range chickens (40.4%) and ducks 15% from rural area surrounding Giza. This probably due to free range poultry is mainly infected via environmental oocysts. Additionally, they like omnivore’s classes, possible engulf raw meat residues30. Also, our results are accepted through other studies which validated the vast majority of virulent Egyptian sheep and poultry isolates are corresponding to type II strain which is the predominant human types31. Moreover, from São Paulo state, Brazil, T. gondii was isolated from tissue homogenates of 16 sheep, where 6 of the 16 isolates killed 100% of infected mice32. In contrast, we could not isolate any of T. gondii tissue cyst from the sero-positive cattle (cows and buffaloes), perhaps due to low susceptibility and higher resistance of those species or owing to incompatibility among the taken tissue samples and the parasite harboring ones.
In the present study, mice viability test of meat producing animal identify potential zoonotic hazard via three virulent strains, I, II and III. But, more one type may be harboring the same human or animal hosts33. However, the higher virulent types usually hidden the biological properties of less virulent ones. Thus, the practice of parasite isolation via mice viability possible results a single strain even several strains were found in the inoculum. So, genotyping is more accurate for strain recognition but mice viability test is essential for morbidity and virulence discovery.
Classically, consumption of undercooked meat has been documented as the major risk factor, followed by dusting oocysts and placental diffusion2. Therefore, many reports have indicated that human toxoplasmosis mainly took place in many localities free from cats34. This is in contrast to farm animals that are usually infected by cohabitation with cats31. Where, intensifying awareness against undercooked meat minimizing human prevalence world-wide. So, the incidence of human toxoplasmosis completely differs from country to other according to the socio-economic pattern of meat feeding behavior. Wherever, various communities cover diverse culture habits are contributed to the zoonotic aspects for spreading toxoplasmosis35.
Insufficient cooked meat is the main risk factor but not the absolute one. Occasionally some strains may survive deep-freezing36. Microwaving or superficial heating does not destroy all bradyzoites and remain viable in refrigerated carcasses (1-4°C) or minced meat for up to 3 weeks37, also survive freezing at temperatures between -1 and -8°C for longer than a week and cyst containing bradyzoites remain viable and infective for as long as the meat is edible38, Accordingly, changing our eating behavior possible minimizes the high incidence of human toxoplasmosis. Where, dormant cysts in meat was rendered non-viable when internal temperatures had reached 67 or -12°C freeze for 1 day35.
CONCLUSION
Bio-viability and molecular recognition along with genotyping of T. gondii Egyptian isolates from human and animal hosts are on target bio-indicators, signify zoonotic impact through virulent strains and must be focused for avoiding feeding on under cooked meat of animals or birds.
SIGNIFICANCE STATEMENTS
This study evaluate the zoonotic hazard of T. gondii seropositive Egyptian animals and aborted women via mice viability test and genotyping, as excellent bio-indicators for virulent strains. However, the microscopic confirmed bradyzoites containing ones were bio-assayed through intra-peritoneal passage in mice as viability test. The total seropositive percentages and the suspected microscopic examined samples plus to DNA percentages were detected. Also, the total succeeded isolates via mice viability test were (6, 8, 4, 0, 0, 2 and 2) corresponding to (women, sheep, goat, cows, buffaloes, camels and free range poultry) respectively. Genotyping confirmed 22 isolates (2 type-I, 13 type-II and 7 type-III). Mice viability and DNA approve along with genotyping were signify T. gondii virulent types I, II and III viable cysts from Egyptian women, animals and poultry and reflect zoonotic bio-hazard of public health concern.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the scientific and financial support of National Research Centre through the Project number P 101202 (2015-2016) entitled: Molecular Detection and Genotyping of Toxoplasma gondii from Egyptian isolates.
REFERENCES
- Innes, E.A., 2010. A brief history and overview of Toxoplasma gondii. Zoonoses Public Health, 57: 1-7.
CrossRefPubMedDirect Link - Tenter, A.M., A.R. Heckeroth and L.M. Weiss, 2000. Toxoplasma gondii: From animals to humans. Int. J. Parasitol., 30: 1217-1258.
CrossRefPubMedDirect Link - Fromont, E.G., B. Riche and M. Rabilloud, 2009. Toxoplasma seroprevalence in a rural population in France: Detection of a household effect. BMC Infect Dis., Vol. 9.
CrossRefDirect Link - Kovari, H., C. Ebnother, A. Schweiger, N. Berther, H. Kuster and H.F. Gunthard, 2010. Pulmonary toxoplasmosis, a rare but severe manifestation of a common opportunistic infection in late HIV presenters: Report of two cases. Infection, 38: 141-144.
CrossRefDirect Link - Bartoszcze, M., K. Krupa and J. Roszkowski, 1991. ELISA for assessing Toxoplasma gondii antibodies in pigs. Zoonoses Public Health, 38: 263-264.
CrossRefDirect Link - Giordano, L.F.C., E.P. Lasmar, E.R.F. Tavora and M.F. Lasmar, 2002. Toxoplasmosis transmitted via kidney allograft: Case report and review. Trans. Proc., 34: 498-499.
CrossRefDirect Link - Wulf, M.W.H., R. van Crevel, R. Portier, C.G. ter Meulen, W.J.G. Melchers, A. van der Ven and J.M.D. Galama, 2005. Toxoplasmosis after Renal transplantation: Implications of a missed diagnosis. J. Clin. Microbiol., 43: 3544-3547.
CrossRefDirect Link - Robert-Gangneux, F. and M.L. Darde, 2012. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev., 25: 264-296.
CrossRefPubMedDirect Link - Elfadaly, H.A., M.A. Hassanain, R.M. Shaapan, A.M. Barakat and N.I. Toaleb, 2012. Serological and hormonal assays of murine materno-fetal Toxoplasma gondii infection with emphasis on virulent strains. World J. Med. Sci., 7: 248-254.
Direct Link - Hassanain, M.A., H.A. El-Fadaly and N.A. Hassanain, 2014. Toxoplasma gondii parasite load elevation in diabetic rats as latent opportunistic character. Ann. Trop. Med. Pubblic Health, 7: 110-115.
Direct Link - Elfadaly, H.A., M.A. Hassanain, R.M. Shaapan, N.A. Hassanain and A.M. Barakat, 2015. Corticosteroids opportunist higher Toxoplasma gondii brain cysts in latent infected mice. Int. J. Zool. Res., 11: 169-176.
CrossRefDirect Link - Howe, D.K., S. Honore, F. Derouin and L.D. Sibley, 1997. Determination of genotypes of Toxoplasma gondii strains isolated from patients with toxoplasmosis. J. Clin. Microbiol., 35: 1411-1414.
Direct Link - Grigg, M.E and J.C. Boothroyd, 2001. Rapid identification of virulent type I strains of the protozoan pathogen Toxoplasma gondii by PCR-restriction fragment length polymorphism analysis at the B1 gene. J. Clin. Microbiol., 39: 398-400.
CrossRefDirect Link - Ajzenberg, D., A.L. Banuls, M. Tibayrenc and M.L. Darde, 2002. Microsatellite analysis of Toxoplasma gondii shows considerable polymorphism structured into two main clonal groups. Int. J. Parasitol., 32: 27-38.
CrossRefDirect Link - Maubon, D., D. Ajzenberg, M.P. Brenier-Pinchart, M.L. Darde and H. Pelloux, 2008. What are the respective host and parasite contributions to toxoplasmosis? Trends Parasitol., 24: 299-303.
CrossRefPubMedDirect Link - Vallochi, A.L., C. Muccioli, M.C. Martins, C. Silveira, R. Belfort Jr. and L.V. Rizzo, 2005. The genotype of Toxoplasma gondii strains causing ocular toxoplasmosis in humans in Brazil. Am. J. Ophthalmol., 139: 350-351.
CrossRefDirect Link - Khan, A., C. Su, M. German, G.A. Storch, D.B. Clifford and L.D. Sibley, 2005. Genotyping of Toxoplasma gondii strains from immunocompromised patients reveals high prevalence of type I strains. J. Clin. Microbiol., 43: 5881-5887.
CrossRefDirect Link - Zhou, P., H. Zhang, R.Q. Lin, D.L. Zhang, H.Q. Song, C. Su and X.Q. Zhu, 2009. Genetic characterization of Toxoplasma gondii isolates from China. Parasitol. Int., 58: 193-195.
CrossRefPubMedDirect Link - Darde, M.L., B. Bouteille and M. Pestre-Alexandre, 1992. Isoenzyme analysis of 35 Toxoplasma gondii isolates and the biological and epidemiological implications. J. Parasitol., 78: 786-794.
CrossRefDirect Link - Dubey, J.P., 2009. Toxoplasmosis in sheep-The last 20 years. Vet. Parasitol., 163: 1-14.
CrossRefPubMedDirect Link - Dubey, J.P., D.H. Graham, E. Dahl, M. Hilali and A. El-Ghaysh et al., 2003. Isolation and molecular characterization of Toxoplasma gondii from chickens and ducks from Egypt. Vet. Parasitol., 114: 89-95.
CrossRefPubMedDirect Link - Barakat, A.M.A., M.M. Abd Elaziz and H.A. Fadaly, 2009. Comparative diagnosis of toxoplasmosis in Egyptian small ruminants by indirect hemagglutination assay and ELISA. Global Veterinaria, 3: 9-14.
Direct Link - Aubert, D., G.T. Maine, I. Villena, J.C. Hunt and L. Howard et al., 2000. Recombinant antigens to detect Toxoplasma gondii-specific immunoglobulin G and immunoglobulin M in human sera by enzyme immunoassay. J. Clin. Microbiol., 38: 1144-1150.
Direct Link - Burg, J.L., C.M. Grover, P. Pouletty and J.C. Boothroyd, 1989. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol., 27: 1787-1792.
PubMedDirect Link - Lehmann, T., C.R. Blackston, S.F. Parmley, J.S. Remington and J.P. Dubey, 2000. Strain typing of Toxoplasma gondii: Comparison of antigen-coding and housekeeping genes. J. Parasitol., 86: 960-971.
CrossRefDirect Link - Dubey, J.P., B. Lopez, M. Alvarez, C. Mendoza and T. Lehmann, 2005. Isolation, tissue distribution and molecular characterization of Toxoplasma gondii from free-range chickens from Guatemala. J. Parasitol., 91: 955-957.
CrossRefDirect Link - Darde, M.L., 2003. Genetic analysis of the diversity in Toxoplasma gondii. Annali dell'Istituto Superiore di Sanit A, 40: 57-63.
PubMedDirect Link - Abdel-Hameed, D.M. and O.M. Hassanein, 2008. Genotyping of Toxoplasma gondii strains from female patients with toxoplasmosis. J. Egypt. Soc. Parasitol., 38: 511-520.
PubMedDirect Link - Cook, A.J.C., R. Holliman, R.E. Gilbert, W. Buffolano and J. Zufferey et al., 2000. Sources of Toxoplasma infection in pregnant women: European multicentre case-control study Commentary: Congenital toxoplasmosis-further thought for food. BMJ, Vol. 321.
CrossRef - Hassanain, M.A., H.A. Elfadaly, R.M. Shaapan, N.A. Hassanain and A.M. Barakat, 2011. Biological assay of Toxoplasma gondii Egyptian mutton isolates. Int. J. Zool. Res., 7: 330-337.
Direct Link - Boyer, K.M., E. Holfels, N. Roizen, C. Swisher and D. Mack et al., 2005. Risk factors for Toxoplasma gondii infection in mothers of infants with congenital toxoplasmosis: Implications for prenatal management and screening. Am. J. Obstet. Gynecol., 192: 564-571.
CrossRefPubMedDirect Link - Aspinall, T.V., D. Marlee, J.E. Hyde and P.F.G. Sims, 2002. Prevalence of Toxoplasma gondii in commercial meat products as monitored by polymerase chain reaction-food for thought? Int. J. Parasitol., 32: 1193-1199.
CrossRefDirect Link - Frenkel, J.K., 1990. Transmission of toxoplasmosis and the role of immunity in limiting transmission and illness. J. Am. Vet. Med. Assoc., 196: 233-240.
PubMedDirect Link - Dubey, J.P., 1988. Long-term persistence of Toxoplasma gondii in tissues of pigs inoculated with T. gondii oocysts and effect of freezing on viability of tissue cysts in pork. Am. J. Vet. Res., 49: 910-913.
PubMedDirect Link - McDonald, J.C., T.W. Gyorkos, B. Alberton, J.D. MacLean, G. Richer and D. Juranek, 1990. An outbreak of toxoplasmosis in pregnant women in northern Quebec. J. Infect. Dis., 161: 769-774.
CrossRefDirect Link - Dubey, J.P. and D.W. Thayer, 1994. Killing of different strains of Toxoplasma gondii tissue cysts by irradiation under defined conditions. J. Parasitol., 80: 764-767.
CrossRefPubMedDirect Link - El-Nawawi, F.A., M.A. Tawfik and R.M. Shaapan, 2008. Methods for inactivation of Toxoplasma gondii cysts in meat and tissues of experimentally infected sheep. Foodborne Pathogen Dis., 5: 687-690.
CrossRefDirect Link