
Ahmad Abdolzadeh
Not Available
Fatemeh Hosseinian
Not Available
Mahnaz Aghdasi
Not Available
Hameidreza Sadgipoor
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 271-276







ABSTRACT
The main aim of this study was the evaluation of the effects of nitrogen sources on growth of the Catharanthus roseus and the concentration of total alkaloids, vincristin and vinblastin. Plants raised in sand culture in greenhouse and irrigated two times per day with Hoagland solution containing nitrate, ammonium and nitrate plus ammonium as the nitrogen source and at total nitrogen concentrations of 2.75, 5.5, 11, 22 and 32 mM. Plants fed with nitrate plus ammonium showed the greatest increase in shoot dry mass whereas the lowest shoots and total dry mass were exhibited in nitrate treatment. There was no significant difference in shoot or root dry mass between plants fed with different nitrogen concentrations at the 5% probability level but the highest total dry mass was observed in 22 mM nitrogen concentration. The highest content of amino acids, proteins, total nitrogen, total alkaloids, vincristin and vinblastin were observed in plant fed with nitrate plus ammonium. Maximal contents of amino acids, total alkaloids, vincristin and vinblastin occurred at a total nitrogen concentration of 11 mM. Significant correlation was found between vincristin and vinblastin content and total alkaloids, protein and nitrogen. Increase in the nitrogen level beyond 11 mM had an antagonistic effect on alkaloid content. The increase in plant mass was not related to alkaloid levels and maximum the alkaloid production occurred at 11 mM nitrate plus ammonium.
PDF Abstract XML References Citation
How to cite this article
Ahmad Abdolzadeh, Fatemeh Hosseinian, Mahnaz Aghdasi and Hameidreza Sadgipoor, 2006. Effects of Nitrogen Sources and Levels on Growth and Alkaloid Content of Periwinkle. Asian Journal of Plant Sciences, 5: 271-276.
DOI: 10.3923/ajps.2006.271.276
URL: https://scialert.net/abstract/?doi=ajps.2006.271.276
DOI: 10.3923/ajps.2006.271.276
URL: https://scialert.net/abstract/?doi=ajps.2006.271.276
INTRODUCTION
As one of the most important essential elements in plants (Kirkby, 1981), the soil nitrogen forms and levels have marked effect on plant growth and development and affecting special metabolic pathways in plants (Cramer and Lewis, 1993; Fabre and Planchon, 2000). However, little is known about the plant nitrogen nutritional statues and its effects on nitrogen containing substances like alkaloids (Bhadra and Shanks, 1997; Garner et al., 1996; Morgan, 2000). Nitrate and ammonium are major forms of inorganic nitrogen taken up by the roots of higher plants. Nitrate is readily mobile in the xylem and can be stored in vacuoles of root and shoot parenchyma cells without toxicity. However, in order to be assimilated and fulfill its essential functions, it has to be reduced to ammonium. Plants are readily able to take up ammonium, especially when environmental conditions are suitable for high photosynthesis rate. In contrast to nitrate, ammonium assimilation has lower energy cost but most of the absorbed ammonium has to be incorporated into organic molecules in the roots to prevent ammonium toxicity (Kaul and Hoffman, 1993). The preferential uptake of ammonium or nitrate is related to several factors including plant species, root medium pH, temperature and nitrogen concentration (Marschner, 1995). On the other hand, soil nitrogen level has marked effect on vegetative and reproductive growth. Nitrogen deficiency leads to growth reduction, early senescence and low shoot to root ratio. On the contrary, excess nitrogen results in promoted vegetative growth and delays plant reproductive growth. It is expected that these processes have an important bearing on special metabolic pathways in plants (Marschner, 1995; Kirkby, 1981).
Periwinkle is an annual and medicinal plant from Apocynaceae family that produces over ninety terpenoid indole alkaloids. The most important of these alkaloids are ajmalicine and serpentine used in the treatment of hypertension and vincristin and vinblastin with an antineuplasy effect (Correia, 2001). Vincristin and vinblastin are dimmer alkaloids that have been derived from strictosidine. The latter is produced through the condensation of tryptamine with secologanin. The tryptamine is the product of tryptophan decarboxylation reaction. In these steps, vindoline and catharanthine are produced first and then combined to form vincristin and vinblastin which accumulate in the vacuoles of leaf cells (Bhadra et al., 1998). The concentrations of these alkaloids are very low in plants and not synthesized to any appreciable extent in cell culture. The reason for this is thought to be tissue-specific expression of alkaloid synthesizing genes, because in some cases plants regenerated from non-producing callus cells contained the same alkaloid profiles as the parent plants (Buchanan et al., 2000). Therefore, attempt should be directed towards finding optimal growth and nutritional conditions which leads to greater alkaloid production in cultured plants. It is reported that nitrate and ammonium ions have different effects on secondary metabolite synthesis. Nitrate is known to promote secondary metabolite synthesis, while ammonium inhibits it. Also, the nitrogen level may affect alkaloid content of the plants (Zhong and Wang, 1998; Hahn et al., 2003). Therefore, it is very important to find optimal nitrogen sources and levels suitable to the plant species for secondary metabolite production.
Accordingly, in this research we examined the effects of nitrogen source and levels on the content of total alkaloids, vincristin, vinblastin, amino acids, proteins and total nitrogen as well as some growth parameters. The main aim of this study was to determine the suitable nitrogen source and level for the highest growth rate and maximal vincristin and vinblastin production.
MATERIALS AND METHODS
All of the experiments were carried out at the Gorgan university of agricultural science and natural resource during 2003-2004. Seeds of periwinkle (Catharanthus roseus L.) were obtained from local market and screened for the uniform size and color. They were germinated in Petri dishes lined with moist filter paper and incubated in dark at 20±1°C. Germinated seeds were subsequently transferred into pots filled with sand. The plants raised in greenhouse and irrigated with Hoagland solution for the first two weeks. Thereafter, they were treated with modified Hoagland based solutions depending on the experiment. In one set of experiments plants were treated with Hoagland solution containing different nitrogen sources at a total nitrogen concentration of 11 mM. The nitrogen sources consisted of nitrate, ammonium and nitrate plus ammonium (NO3-/NH4+ ratio 1/1.2). In the second set of experiments five nitrogen levels (2.75, 5.5, 11, 22 and 32 mM) were used in the form of a nitrate plus ammonium where the ratio of NO3-/NH4+ ratio were kept at 1/1.2. The pH of the nutrient solution was adjusted daily at 6.2±0.2 and nutrient solutions were changed every week. The averages of daily maximum and minimum temperatures in greenhouse during the growing period (spring of 2003) were 30 and 17°C, respectively. Relative humidity was between 55 and 75%. Plants harvested after four months. Total nitrogen was quantified after digestion with H2SO4 by Kjeldahl methods using Micro Kjeldahl Tecator Model 103 (Gupta, 1999). Total amino acid was determined after extraction of plant tissue with sulphosalsilic acid using the method described by Yemm and Cocking (1954). Total protein was determined by the method of Lowry et al. (1951) following the extraction of plant tissue with 0.2 M phosphate buffer pH 6.8. The extraction of total alkaloids was carried out as described in Monforte-González et al. (1992). Three gram powders prepared from dried leaves were mixed with 20 cm3 ammonium solution. After 20 min 30 cm3 chloroform were added and the resulting solution stirred for 2.5 h and then filtered with filter paper (Whatman No. 1). The filtrate solution was concentrated to 15 cm3 by evaporation in water bath at 30°C and mixed with 15 cm3 of 2% (w/v) tartaric acid. The aqueous phase was separated in a decantor and the pH of the solution adjusted to 9 with the ammonium solution. Alkaloids were solubilized in 10 mL glacial acetic acid. Total alkaloid in the acetic acid solution was determined after titration with 0.5 N perchloric acid using Crystal Violet as an indicator. Vincristin and vinblastin were determined after separation of methanol extracted alkaloids with thin layer chromatography (Silicagel G60-UV254, Merk) according to the method of Monforte-González et al. (1992). The separated TLC bands were identified by dragendorf reagent. Bands were scrapped from the TLC plate, dissolved in methanol and analyzed spectrophotometrically for vincristin and vinblastin. Their absorption spectra in the range of 200-500 nm were recorded using U.V. spectrophotometer (Shimadzo Japan) (Kutney et al., 1980).
All the statistical analysis were performed using SAS statistical software. Comparisons of means were performed using LSD test.
RESULTS
Growth: There were no significant differences between either growth parameters under different nitrogen sources and levels at the 5% probability level (Table 1 and 2). The best nitrogen sources for growth were ammonium plus nitrate where total dry mass was significantly higher than ammonium or nitrate fed plants at the 10% probability level. The greatest increase in growth parameters occurred at 22 mM nitrogen that was significantly higher than other nitrogen levels at the 10% probability level. No significant differences were observed in shoot/ root ratio under different nitrogen sources and levels however, this ratio increased slightly by increment of nitrogen levels.
| Table 1: | Effects of different nitrogen sources (totally 11 mM N concentration) on growth factors (g) of periwinkle after 4 months treatments |
 | |
| Means followed by the same letter(s) are not significantly different at the 5% probability level using LSD | |
| Table 2: | Effects of different nitrogen levels as nitrate plus ammonium (1/1.2 ratio) on growth factors (g) of periwinkle after 4 months treatments |
 | |
| Means followed by the same letter(s) are not significantly different at the 5% probability level using LSD | |
| Table 3: | Effects of different nitrogen sources (totally 11 mM N concentration) on concentration of amino acids (μg/g dw), protein (μg/g fw), total N (mg/g dw), total alkaloids (mg/g dw), vincristin and vinblastin (μg/g dw) after 4 months treatments |
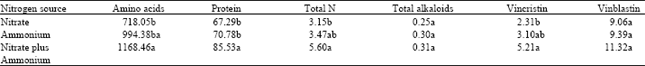 | |
| Means followed by the same letter(s) are not significantly different at the 5% probability level using LSD | |
| Table 4: | Effect of different nitrogen levels as nitrate plus ammonium (1/1.2 ratio) on amino acids (μg/g dw), protein (μg/g fw), total N (mg/g dw), total alkaloids (mg/g dw), vincristin and vinblastin (μg/g dw) concentration after 4 months treatments |
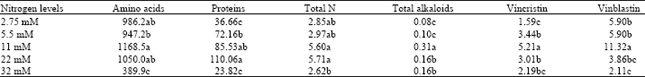 | |
| Means followed by the same letters are not significantly different at the 5% probability level using LSD | |
| Table 5: | The correlation coefficient of measured parameters in different nitrogen source treatments |
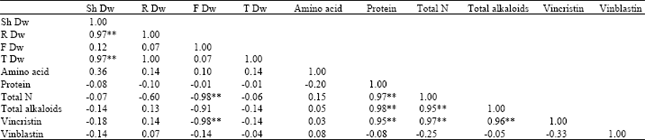 | |
| Note. * and ** significant at 0.05 and 0.001 probability, respectively | |
Effects of nitrogen sources on some biochemical aspects: The highest amounts of total amino acid, protein and nitrogen were observed in plants fed with nitrate plus ammonium (Table 3). However, the lowest amino acid, protein and total nitrogen contents were exhibited in nitrate- fed plants. There were no significant differences in total alkaloid content between different nitrogen source treatments. But the highest content of vincristin and vinblastin was observed in nitrate plus ammonium treatment.
Significant negative correlation was found between vincristin and total nitrogen content with flower dry mass (Table 5). Beginning of the reproductive stage reduces the content of these chemical in leaves. Also, the amounts of vincristin correlated significantly with the total content of alkaloid, protein and nitrogen. However, no significant correlation was found between either the content of vinblastin and flower dry mass or content of vinblastin and amounts of protein and total nitrogen.
Effects of nitrogen levels on some biochemical aspects: The least amounts of total amino acid, protein and nitrogen were observed in plants growing at 32 mM nitrogen level (Table 4). On the contrary, the greatest amount of amino acid occurred at 11 mM nitrogen and the highest protein and total nitrogen contents were at 22 mM nitrogen treatments.
The lowest total alkaloid and vincristin contents were found in plants grown at 2.75 mM nitrogen although, the lowest vinblastin content occurred at 32 mM nitrogen level (Table 4).
| Table 6: | The correlation coefficient of measured parameters in different nitrogen levels treatments |
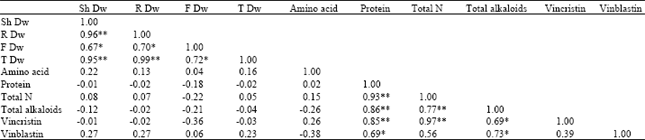 | |
| Note * and ** significant() at 0.05 and 0.001 probability respectively | |
In contrast, the greatest total alkaloids, vincristin and vinblastin contents were observed at 11 mM nitrogen treatment.
There was significant correlation between vincristin content with total alkaloid, protein and nitrogen (Table 6). No significant positive correlation was found between vincristin content with flower dry mass. Also, vinblastin content was positively correlated to the total alkaloid and protein contents.
DISCUSSION
The main objective of this research was to have an evaluation on the best nitrogen source and levels suitable for maximum growth and alkaloid production in periwinkle. As already mentioned, the amount of alkaloids such as vincristin and vinblastin are very low in plants and not synthesized to any appreciable extent in cell culture. Also, there is no established protocol for the industrial production of these important alkaloids with antineuplasy effect (Correia, 2001; Morgan, 2000; Shanks et al., 1998). Given that the supplied nitrogen forms either nitrate or ammonium can have different effects on secondary metabolism. Our study planned to increase the plant endogenous alkaloid levels by manipulating nitrogen status of the culture medium. The best nitrogen sources for the highest growth were ammonium plus nitrate at 22 mM nitrogen. Bialczyk et al. (2004) in tomato, Kotsiras et al. (2005) in cucumbers, Ciesiołka et al. (2005) in Lupinus albus and Zhang et al. (2005) in spinach reported the higher growth and biomass production in nitrate plus ammonium fed plants in compare with sole ammonium or nitrate fed ones. Also, increment of plant growth by increasing of nitrogen levels until special concentration has been reported by several investigators (Marschner, 1995; Al-Humaid, 2004).
There was significant correlation in the amounts of vincristin and total alkaloid with protein and nitrogen contents. These results indicate higher protein and total nitrogen content in the plant may lead to the promotion of metabolic pathways involving in the formation of alkaloids. On the contrary, negative correlation in the amount of vincristin, total alkaloids and total nitrogen with flower dry mass in nitrogen source experiments suggests alterations in plant source-sink relationships after the start of flowering. Flowers are strong sink and probably higher translocation of N compounds and carbon skeleton through phloem to flowers reduced N compounds including alkaloids in leaves (Marschner, 1995).
When ammonium or nitrate was used as a sole nitrogen source, plant amino acid, protein and total nitrogen contents decreased and it resulted in the decline of total alkaloid, vincristin and vinblastin contents. This can explained by the finding that ammonium is mainly is fixed by GS/GOGAT pathway into amino acids (glutamine/glutamate) in roots and the resulting decline in the root carbohydrate content probably will diminish further ammonium uptake and assimilation (Marschner, 1995; Tischner, 2000). The nitrate assimilation pathway starts with nitrate uptake followed by nitrate reduction resulting ammonium which is fixed into amino acids. Although, nitrate is reduced in leaves and the required energy is directly provided by photosynthesis, nitrate uptake and assimilation has higher energy demands than ammonium that may further limit its uptake and assimilation (Tischner, 2000). Consequently, plants fed with ammonium or nitrate as a sole nitrogen source indicated the lower amino acid, protein, total nitrogen and alkaloids contents in compare with ammonium plus nitrate fed plants. Plants fed with the combination of ammonium plus nitrate showed maximum total alkaloid, vincristin and vinblastin accumulation. Probably, fine regulation of nitrogen assimilation in the root and shoot, lower energy demands for nitrogen reduction and better charge balance Intracellular pH regulation when supplied simultaneously as nitrate and ammonium lead to higher amino acid, protein and total nitrogen contents in the plants (Marschner, 1995; Allen and Raven, 1987). Present results are consistent with the previous findings by Hahn et al. (2003) who reported inhibition of ginsenoside accumulation in Panax ginseng by ammonium in the culture medium. The greatest adventitious root biomass was obtained at a 1:2 ammonium to nitrate, which also resulted in high ginsenoside productivity. Similar results were obtained in Panax quinquefolium cell culture where ammonium was unfavorable to metabolite formation (Zhong and Wang, 1998). Also, in the cell culture of Litospermum erythrorhizon, shikonin synthesis was inhibited by ammonium ions in the culture medium and the addition of nitrate was required at the end of growth phase (Hahn et al., 2003).
Effects of nitrogen levels on alkaloids content of plants were more severe than nitrogen sources. The higher amino acid, protein and total nitrogen contents and consequently the higher accumulation of total alkaloids, vincristin and vinblastin was observed in plants grown at 11 mM nitrogen level. The lower nitrogen levels (2.75 and 5.5 mM) resulted in lower amino acid, protein and total nitrogen contents and consequently reduced accumulation of total alkaloids, vincristin and vinblastin. Also, at higher nitrogen levels (32 mM) the accumulation of all nitrogen containing compounds including alkaloids were decreased. Sreevalli et al. (2004) has been reported total alkaloid increase in leaves and roots of periwinkle genotypes by N fertilization up to 150 kg ha-1. The increase in contents of leaf and root alkaloids, due to N application, paralleled the increase in leaf and root yields. Similarly, Zhong and Wang (1998) have been reported that the maximum production of ginseng saponin and polysaccharide obtained at 40 mM nitrogen level and both 20 and 80 mM nitrogen resulted lower ginseng saponin and polysaccharide production by cell cultures. Also, total alkaloid and drug (hyoscyamine and scopolamine) contents of datura plants increased with increasing the complex fertilization level to a peak value at 600 kg ha-1. It then, decreased at 800 kg ha-1 level. (Al-Humaid, 2004). It appears that there is optimal nitrogen level for maximum alkaloid accumulation in plants and the higher and lower nitrogen levels could result in the lower alkaloids content.
The results indicated that the best nitrogen source for maximum alkaloids production is nitrate plus ammonium. Increasing nitrogen level in the root medium up to a special value led to increment of alkaloids content in plants. However, further N fertilization reduced alkaloids accumulation. Additional studies are required to confirm these results in the field. Since the content of alkaloids in some plants has been found to be influenced by individual factors, such as stage of plant growth, moisture stress (Hahn et al., 2003) and N fertilization, further experiments are needed to determine the combined influence of these factors for maximizing the alkaloid content.
REFERENCES
- Bhadra, R. and J.V. Shanks, 1997. Transient studies of nutrient uptake, growth and indol alkaloids accumulation in heterotrophic cultures of hairy roots of Catharanthus roseus. Biotechnol. Bioeng., 55: 527-534.
CrossRefDirect Link - Ciesiolka, D., M. Muzquiz, C. Burbano, P. Altares and M.M. Pedrosa et al., 2005. An Effect of various nitrogen forms used as fertilizer on Lupinus albus L. yield and protein, alkaloid and a-galactosides content. J. Agron. Crop Sci., 191: 458-463.
Direct Link - Correia, J.J., 2001. Physiochemical aspects of tubulin-interacting antimitotic drugs. Curr. Pharm. Design, 7: 1213-1228.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Monforte-Gonzalez, M., T. Ayora-Talavera, I.E. Maldonado-Mendoza and V.M. Loyola-Vargas, 1992. Quantitative analysis of serpentine and ajmalicine in plant tissues of Catharanthus roseus and hyoscyamine and scopolamine in root tissues of Datura stramonium by densitometry in thin layer chromatography. Phytochem. Anal., 3: 117-121.
- Sreevalli, Y., R.N. Kulkarni K. Baskaran and R.S. Chandrashekara, 2004. Increasing the content of leaf and root alkaloids of high alkaloid content mutants of periwinkle through nitrogen fertilization. Ind. Crops Prod., 19: 191-195.
CrossRefDirect Link - Tischner, R., 2000. Nitrate uptake and reduction inhigher and lower plants. Plant Cell Environ., 23: 1005-1024.
Direct Link