
J.B. Endan
Not Available
J.H. Young
Not Available
M.A. Awal
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 281-286







ABSTRACT
Two growth chamber experiments were conducted to evaluate the effect of defoliation on the growth and development of peanuts. In Experiment one, peanuts (Arachis hypogaea L., cv. Flogiant) were uniformly defoliated by hand at 25, 50, 75 and 100% at the pod or the pod-filling stage. In experiment two, 25 and 75% of the young leaves were removed by hand at the vegetative or the pod-performing stage. Results of these study showed that defoliation lowered the leaf, stem, root, peg and pod masses. The growth stage of the plants determines the variables that would be most affected. The reduction in mass due to defoliation lasted for four to six weeks when defoliation were at the pod-forming or the pod-filling stage and about two to three weeks when 25% of the young leaves were removed at the vegetative stage. Plants defoliated at the 75% level gave greater priority for peg and pod growth than plants defoliated at the 25% level. Defoliated plants have the lighter pods at harvest.
PDF Abstract XML References Citation
How to cite this article
J.B. Endan, J.H. Young and M.A. Awal, 2006. Effect of Hand Defoliation on Peanut Growth. Asian Journal of Plant Sciences, 5: 281-286.
DOI: 10.3923/ajps.2006.281.286
URL: https://scialert.net/abstract/?doi=ajps.2006.281.286
DOI: 10.3923/ajps.2006.281.286
URL: https://scialert.net/abstract/?doi=ajps.2006.281.286
INTRODUCTION
Modeling of peanut growth and development as well as yield prediction with crop simulation models is an important part of peanut research (Ferreyra et al., 2000). Stability of parameters describing crop growth of peanut is important because of the diversity of climatic conditions in which peanuts are grown and is valuable when developing simulation models for this species (Kinirya et al., 2005). Defoliation is an important considering parameter for peanut growth model. Studies on defoliation of peanuts have suggested that severe defoliation after the early vegetative stage affect yields (Campbell, 1978; Santos and Sutton, 1983; Turner, 1982; Wilkerson et al., 1984). Most defoliation studies in the literature were concerned with yields. Only one study (Turner, 1982) was intended for modelling insect defoliation of peanuts.
Result from the previous studies on peanuts growth (Cox, 1978; 1979) at the phototonus unit of the South eastern Plant Environment Laboratory at North Carolina State University were successfully used to develop a peanut growth model, PEANUT (Young et al., 1979). The model accounts for this loss by removing an appropriate portion of then leaf mass at the desired time Simulation of defoliation using PEANUT have given valuable information on the effect of defoliation on growth and development of peanuts. The model predicted trends of yield reduction similar to those observed in studies made by Campbell (1978), with the critical time for defoliation occurring at pod-forming and pod-filling stages. However, there were substantial differences between yields predicted by the model and those with experiments. The reason for the discrepancies were unclear, since there was inadequate information to ensure correct input of all soil and weather condition in the model. Nevertheless, some model modification may be required since corn earworm prefer to feed on young peanuts leaves, new shot growth and possibly on flowers and pegs (Smith and Barfield, 1982). Corn earworm will proceed to feed on older leaves as they grow and as their food requirement become greater than new shoot growth can provide. Hence, defoliation is a progressive even rather than a one-time removal of leaves as depicted by the present model. The objectives of this study were to compare effects of uniform defoliation with those of removal of young leaves on growth and development of peanuts, Arachis hypogaea L., cv. Florigiant and to study the effect of timing of defoliation on growth and development of Florigiant peanuts.
MATERIALS AND METHODS
This study was conducted at Southeastern Plant Environment Laboratory, North Carolina State University, Releigh in 1988. Two types of defoliation performed in this study were:
Experiment 1
| • | Uniform defoliation by hand at the pod-forming stage (day 70 of growth), |
| • | Uniform defoliation by hand at the pod-filling stage (day 91 of growth), |
Experiment 2
| • | Defoliation of youngest leaves by hand at the vegetative stage (day 35 of growth), |
| • | Defoliation of youngest leaves by hand at the pod-forming stage (day 70 of growth). |
In Experiment 1, Florigiant peanuts were planted at a rate of 4 seeds per pot in 25 cm pots filled with 1/2 peat 1/2 sand. Plants were grown in two walk-in chambers identically set at 30/26°C day/night temperatures and 15 h of photosynthetically active radiation at 680 μE m-2 S-1. Seedlings were thinned to one per pot within 10 days. Pots were placed on carts arranged in three rows of six carts per row in each chamber. Each cart held four pots, giving a density of 8.3 plants m-2 in the two chambers. Plants were irrigated twice daily and supplied with complete nutrient solution twice each week. On day 70 or day 91°C pod-forming or pod-filling stage respectively, ie., stage R3 and R5 as described by Boote (1982), some plants were defoliated uniformly at 25, 50, 75 or 100% by removing one, two, three or all leaflets from each leaf on each plant. Four plants (two from each chamber) were destructively sampled every week for the control and two plants each (one from each chamber) for the four levels of defoliation were sampled weekly after day 70 and day 91. Flower counts; masses of leaves, stems, roots, pegs and pods; and numbers of pegs and pods were recorded. Carts were moved weekly and randomly locked together after each flower count so as to minimize effect of differences in air circulation and light levels within the chamber.
In Experiment 2, Florigiant peanuts were planted in 11 cm diameter pots filled with 1/2 peat 1/2 sand. Two seeds were sawed in each pot and seedlings thinned to one after 10 days and transplanted into 25 cm pots (Experiment 2a) or plastic trays (Experiment 2b) measuring 50x35x10 cm at two seedlings per tray. Each tray was placed on a cart arranged in two rows of six carts per row for each chamber giving a plant density of 2.7 plants m-2. The two chambers were maintained as in the first experiment. For this 12 study, 25 or 75% of the young leaves were removed on day 35 (vegetative growth stage) or day 70 (pod-forming stage). Levels of defoliation were achieved by removal of the desired numbers of younger leaves from each lateral or sub-lateral after counting the total number of leaves (A defoliation table was constructed to assist in determining the number of leaflets that had to be removed). Masses of leaves, stems, roots, pegs and pods were taken weekly for two weeks for plants defoliated on day 35 and once every two weeks for 8 weeks for plants defoliated on day 70. For each defoliation treatment, four plants (two plants per tray per chamber) were destructively sampled on a particular day.
RESULTS AND DISCUSSION
Defoliation affected almost every plant component (leaf, stem, root, peg and pod). The reduction in plant mass was evident for about two weeks when young leaves were removed at the 25% level during the vegetative stage and for about four to six weeks when young leaves were removed at the pod-forming stage or when uniform defoliations were performed at pod-forming or pod-filling stages (Fig. 1). After this stress period, defoliated plants increased growth. Plants that were defoliated at the 100% level took longer to recover (Fig. 1A and B) and (2A and B). The growth stage determined the variables affected by the defoliation (Table 1-3). Plants defoliated uniformly at pod-forming stage (Table 1) had significantly lower leaf number, leaf area, leaf mass, stem mass, peg number and peg mass for at least four weeks after defoliation. Root mass, number of pods and pod mass were not significantly different between the control and the defoliated plants. Plants uniformly defoliated at pod-filling stage (Table 1) had significantly lower leaf number, leaf area, leaf mass and peg mass for as long as four weeks after defoliation.
Uniform defoliation at the pod-forming stage caused greater reduction than uniform defoliation at the pod-filling stage. This is shown by the greater % reduction of leaflet number, leaf area, stem mass, peg number, peg mass, pod number and pod mass for the uniform defoliation on day 70 when compared with day 91.
As shown in Table 1, pegs and pods were greatly affected by uniform defoliation at pod-forming when compared with % reduction for uniform defoliation at pod-filling. Reduction in peg mass caused by uniform defoliation at pod-forming was significant and about twice as much as that resulting from uniform defoliation at pod-filling. Pod mass reduction was much greater for day 70 than for day 91 even though differences were not statically significant. Uniform defoliation at pod-forming had more detrimental effects than uniform defoliation at pod-filling.
Removal of young leaves at the vegetative growth stage (Table 2) resulted in a significant lowering of leaf area, leaf mass, stem mass, root mass, peg number and peg mass, but leaf number, pod number and pod mass were not significantly different from the control plants.
| Table 1: | Mean of the control and % reduction with defoliation when plants were defoliated uniformly at the pod forming or pod-filling stage (Exp. la and 1b) |
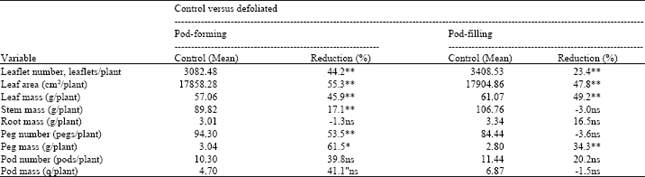 | |
| **Highly significant (α = 0.01), *significant (α = 0.05), ns is not significant. Analysis was based on average of weekly samples for 5 weeks and 4 weeks after defoliation, respectively, for the 2 defoliation timings (day 70 and day 91) | |
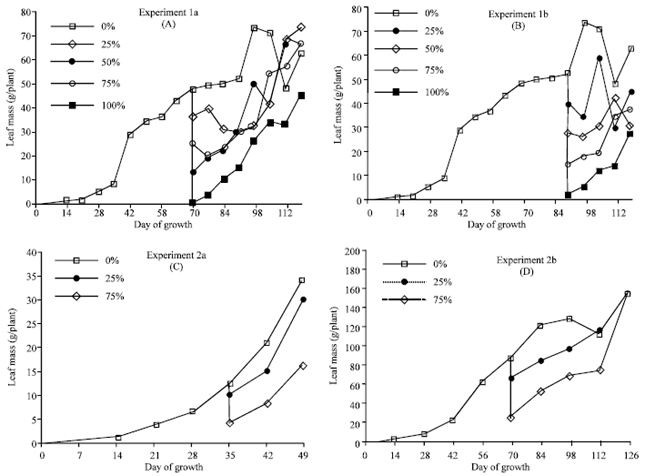 | |
| Fig. 1: | Effect of uniform defoliation (Exp. 1a and 1b) and defoliation of young leaves (Exp. 2a and 2b) on leaf growth |
Plants that had young leaves removed at pod-forming stage (Table 2) had significantly lower leaf number, leaf area, leaf mass, peg number, peg mass, pod number and pod mass. With the exception of the stem and root, removal of young leaves at the pod-forming stage resulted in a greater reduction of the plant components than defoliation at the vegetative growth stage. Therefore, removal of young leaves at the pod-forming stage may be more detrimental to peanuts than similar removal at the vegetative stage.
Table 3 compares uniform defoliation with removal of young leaves at pod-forming. The % reduction for uniform defoliation was calculated from only the 25 and 75% defoliation so as to allow comparison of the reductions between these two methods of defoliation.
| Table 2: | Mean of the control and % reduction with defoliation between control and defoliated plants when young leaves were removed at the vegetative or the pod-forming stage (Exp. 2a and 2b) |
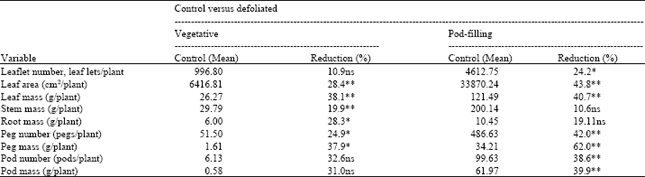 | |
| ** Highly significant (α = 0.01), *significant (α = 0.05), ns is not significant. Analysis was based on average weekly samples for 2 weeks after defoliation and once every 2 weeks for 4 weeks for defoliation on day 35 and 70, respectively | |
| Table 3: | Mean % reduction with defoliation (control versus defoliated) for uniform and defoliation of young leaves at the pod-forming stage (day 70) |
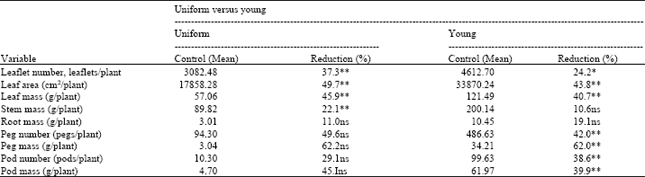 | |
| ** Highly significant (α = -0.01), * significant (α = 0.05), ns is not significant. Analysis was based on average weekly samples for 5 weeks after defoliation and, once every 2 weeks for 4 weeks after defoliation, respectively. Only the control, 25 and 75% samples were used for this analysis | |
| Table 4: | Mean of the 25% and % difference of the 75% from the 25% defoliated plants when young leaves were removed at the vegetative or the pod-forming stage (Exp. 2a and 2b) |
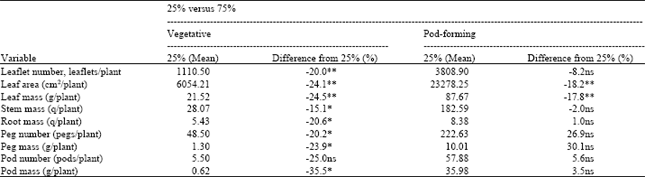 | |
| **Highly significant (α = 0. 01) ' significant (α =0. 05); ns is not significant. Analysis was based on average weekly samples for 2 weeks after defoliation and once every 2 weeks for 4 weeks after defoliation on Day 35 and 70, respectively | |
From Table 3, note that uniform defoliation affected the leaves more than removal of young leaves. Even though % defoliation levels were the same (based on the number of leaves) the % reduction in leaf mass for the removal of young leaves was less than that for uniform defoliation (24.2 versus 37.3%). This was probably due to greater and speedier defoliation with removal of the young shoots. The differences between the % peg and pod reductions for the two methods of defoliation were not conclusive. Loss of leaf mass with the removal of young leaves was less than for uniform defoliation at the same defoliation levels because of the higher specific number (number of leaves per gram) of young leaves. This may also be the reason for the greater % reduction in leaf growth for the uniform defoliation.
From Table 4, note that plants defoliated at 75% during the vegetative growth stage had smaller leaflet number, leaf area, leaf mass, stem mass, root mass, peg number, peg mass, pod number and pod mass than those defoliated at the 25% level.
| Table 5: | Mean of the 25% defoliated plants and % difference of 75% from the 25% when plants were defoliated uniformly at the pod-forming or pod-filling stage (Exp. la and 1b) |
 | |
| ** Highly significant (α = 0.01), * significant (α = 0.05), ns is not significant. Analysis was based on average weekly samples for 5 weeks and 4 weeks after defoliation for 2 defoliation timings (day 70 and day 91), respectively | |
These differences were substantial and consistent, indicating that differences between 75 and 25% defoliation on plant components were about the same (approximately 23% reduction) when plants were defoliated at the vegetative growth stage. However, 75% removal of young leaves at pod-forming gave higher root mass, peg number, peg mass, pod number and pod mass than 25% removal of young leaves. Differences were not consistent, suggesting that there was priority in the partitioning of photosynthetic to various plant parts. Since 75% defoliation resulted in greater reproductive growth (peg and pod growth), the 75% defoliated plants must have given more priority for peg and pod growth than the 25% defoliated plants. Table 5 shows the mean differences between 25 and 75% defoliation at the pod-forming and the pod-filling stage. Uniform defoliation at 75% during the pod-forming stage produced lower leaf, peg and pod values than 25% defoliation. However, at the pod-filling stage, the 75% defoliated plants showed increased peg and pod growth, thus, indicating that the 75% defoliated plants gave more priority to peg and pod growth than the 25% defoliated plants. Similarly, removal of young leaves at the 75% level during pod-forming resulted in higher peg and pod growth (Table 4). Therefore, the effect of uniform defoliation at pod forming was different from the effect of removal of young leaves at the same stage.
Specific leaf area increased with growth. When plants were defoliated, some of the stored reserve material in the stems was probably redirected to the shoot region for new growth; otherwise, the stem may have played an active role in the manufacture of photosynthetic for new shoot growth. This was shown by the fact that plants that were defoliated at the 100% level did not die. Since the analysis of results for defoliation on day 70 did not show that stem mass was significantly reduced, amount of photosynthetic required for new shoot growth was probably very small when compared with the stem mass of mature plants. Root mass of defoliated plants was not different from the control in all cases except when plants were defoliated at the vegetative stage. Root mass formed a major component of the total plant mass, when plants were 35 days old. Defoliation at this stage significantly affected the roots. As the plants grew, leaf, stem, peg and pod growth became major events, exceeding root growth resulting in the roots becoming only a small portion of the total plant mass.
Peanut plants were able to recover and continue growth following various levels of defoliation. Growth stages of the plants determined the plant components that were most affected by defoliation. Uniform defoliation at pod-forming had a more detrimental effect than uniform defoliation at pod-filling stage. Removal of young leaves at the pod-forming stage was more detrimental than removal of young leaves at the vegetative stage. Removal of young leaves at pod-forming resulted in speedier defoliation and recovery than uniform defoliation at the same level suggesting that uniform defoliation may be more detrimental to peanuts than removal of young leaves. Plants defoliated at the 75% level gave greater priority for peg and pod growth than plants defoliated at the 25% level. When plants were completely defoliated, some stored reserve material in the stem was probably redirected to the shoot region for new growth; otherwise, the stem may have played an active role in the manufacture of photosynthetic for shoot growth. Pod formation and growth were delayed in defoliated plants resulting in smaller pods at harvest.
REFERENCES
- Ferreyra, R.A., L.B. Pachepsky, D. Collino and B. Acock, 2000. Modeling peanut leaf gas exchange for the calibration of crop models for different cultivars. Ecol. Model., 131: 285-298.
Direct Link - Kinirya, J.R., C.E. Simpsonb, A.M. Schubertc and J.D. Reedc, 2005. Peanut leaf area index, light interception, radiation use efficiency and harvest index at three sites in Texas. Field Crops Res., 91: 297-306.
Direct Link - Santos, R.B. and B.G. Sutton, 1983. Effect of defoliation on the reproductive development of the peanut. Aust. J. Agric. Res., 34: 527-535.
Direct Link - Wilkerson, G.G., J.W. Jones and S.L. Poe, 1984. Effect of defoliation on peanut plant growth. Crop Sci., 24: 526-531.
Direct Link