
J.W. Muthomi
Not Available
J.K. Ndung`u
Not Available
G.N. Chemining`wa
Not Available
J.M. Wagacha
Not Available
Asian Journal of Plant Sciences
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 585-591







ABSTRACT
Eight cultivars of wheat that are commonly grown in Nakuru and Nyandarua districts of Kenya were tested for their susceptibility to Fusarium head blight (scab) under green house conditions. The cultivars were inoculated with mixed inoculum derived from three pathogenic isolates of Fusarium graminearum that had previously been isolated from wheat. Head blight severity was assessed using a 1-9 scale based on proportion of spikelets bleached and the area under disease progress curve was derived from the disease severity data. At harvest, kernel weight reduction as compared to the untreated controls was determined. All the 8 wheat cultivars were found to be susceptible but they differed in the level of susceptibility. Disease severity among the cultivars varied from 5-59% while the area under disease progress curve varied from 93-994. Cultivars ‘mbuni’ and ‘chiriku’ were the most susceptible, with a grain weight reduction of up to 74%. The results indicate that most of the varieties grown in Nakuru and Nyandarua districts are susceptible to Fusarium head blight. The study indicated that Njoro Bw1 and Njoro Bw2 are the most promising. The resistance trait in these varieties could be useful in Fusarium head blight management.
PDF Abstract XML References Citation
How to cite this article
J.W. Muthomi, J.K. Ndung`u, G.N. Chemining`wa and J.M. Wagacha, 2007. Reaction of Some Kenyan Wheat Cultivars to Head Blight after Inoculation with Fusarium graminearum
. Asian Journal of Plant Sciences, 6: 585-591.
DOI: 10.3923/ajps.2007.585.591
URL: https://scialert.net/abstract/?doi=ajps.2007.585.591
DOI: 10.3923/ajps.2007.585.591
URL: https://scialert.net/abstract/?doi=ajps.2007.585.591
INTRODUCTION
Fusarium Head Blight (FHB) or (scab) of wheat is a common disease of wheat, barley and oats and can result in a reduction in grain quality and yield (Parry et al., 1995). The significance of the disease in wheat production is attributed to both yield reduction and mycotoxin contamination of the grain harvested from the infected ears. The disease is caused by different Fusarium species, including F. graminearum, F. culmorum, F. avenaceum, F. poae, F. crookwellense, F. sporotrichioides and Macrodochium nivale (Parry et al., 1995). Fusarium graminearum and F. culmorum are the most virulent, causing severe blighting of wheat ears (Manka et al., 1985; Stack and McMullen, 1985). Several outbreaks of severe on wheat, mainly associated with F. graminearum, have been experienced since 1990 in different parts of the world. Economic losses in USA were estimated at over $2.5 billion during 1992-1998 (Windels, 2000) and $2.7 billion in 1998-2000 (Nganje et al., 2004). Fusarium graminearum also infects maize causing pink ear rot (Sutton, 1982). The fungi can infect all the vegetative and reproductive parts of cereal crops and survives in crop residues (Jones, 2000).
Fusarium species that infect cereals inhabit soils and are capable of surviving saprophytically on crop debris (Jones, 2000; Parry et al., 1995) and ploughing to burry crop debris removes the source of inoculum from the soil surface which could be available for dispersal to ears. Pathogen population in wheat field soils fluctuate throughout the season, increasing during dry conditions that favour pathogen activity on stem bases (Goswami and Kistler, 2004; Bateman and Murry, 2001; Bateman et al., 1998; Vigier et al., 1997). Conidia are dispersed either by wind or rain splash to wheat heads (Fernado et al., 1997; Jenkinson and Parry, 1994). There is no evidence that seedborne inoculum contributes to Fusarium head blight (Bateman, 2005) but the pathogen may be dispersed by seed (Mishraa et al., 2002). The conidia infect ears mainly during anthesis (Bai and Shanner, 1996). The success of infection depends on weather conditions such as temperature (Brennan et al., 2005; Cowger, 2005; Stein et al., 2005; De Wolf, 2003; Mentewab et al., 2000), humidity (Cowger, 2005; Nita et al., 2005), cultivar resistance (Cowger, 2005; Nita et al., 2005; Llorens et al., 2004) and nitrogen fertilization (Doohan et al., 2003). However, it is the availability of moisture that is the overriding factor (Lacey et al., 1999).
Management options include cultural practices, use of fungicides, breeding for resistance and biological control. Cultural management options include crop rotation, appropriate use of fertilizers, irrigation, weed control, proper land preparation and timely harvesting. Minimum tillage and continuous cereal cropping increases the incidence and severity of Fusarium head blight (Pereyra and Dill-Macky, 2004). It is critical to avoid planting wheat adjacent to fields with large amounts of small grain or maize residues remaining on the soil surface. No till planting of wheat into maize residues substantially increases Fusarium head blight infection. Rotation with a legume crop between maize and small grain crops provides time for the residues to break down and the pathogen population to decline (Champeil et al., 2004). Control of Fusarium head blight using fungicides has been reported to give inconsistent results due to the complexity of the causal organism, influence of nitrogen fertilization, timing of fungicide application and masking control of one Fusarium species by the subsequent growth of another species (Heier et al., 2005; Ramirez et al., 2004; Magan et al., 2002; McMullen et al., 1997; Parry et al., 1995). Application of some fungicides has been shown to stimulate deoxynivalenol and nivalenol production particularly at sub-optimal fungal growth conditions and low fungicide dosage (Ramirez et al., 2004; Magan et al., 2002; Jennings et al., 2000; D’Mello et al., 1999). In addition, food safety concerns limit the chemical management options due to fungicide residues in grain and wheat products (Magan and Olsen, 2004; Jones, 2000).
Most wheat cultivars available are susceptible to Fusarium head blight, none are immune and only a few are moderately resistant (Muthomi et al., 2002a; Parry et al., 1995). However, the use of resistant cultivars still remains the most effective, environmentally friendly, economical and safe strategy of containing the disease and associated mycotoxin contamination. Significant levels of partial resistance that limit yield loss and mycotoxin accumulation have been reported (Pereyra and Dill-Macky, 2004; Wisniewska and Kowalczk, 2005; Miedaner, 1997). Sources of resistance have been found in China, South America and Czech Republic (Lin et al., 2006; Mesterhazy et al., 1999; Mesterhazy, 1995). Wheat varieties exhibit non-specific resistance to head blight due to the large number of Fusarium species involved and lack of genes for complete resistance.
In this study, 8 wheat varieties commonly grown in Nyandarua and Nakuru Districts of Kenya were challenged with Fusarium graminearum under green house conditions with the objective of determining their susceptibility to Fusarium head blight based on symptom expression and yield reduction.
MATERIALS AND METHODS
Survey and plant cultivation: A total of 90 farms were surveyed in Nakuru and Nyandarua districts of Kenya for wheat cultivars grown during the 2004 growing season. The farmers were randomly selected to cover 50 farms in Nakuru and 40 farms in Nyandarua district. For the greenhouse inoculation experiment, certified seeds of 8 wheat varieties (Kwale, Mbuni, Njoro Bw1, Njoro Bw2, Heroe, Chiriku, Chozi and Yombi) were acquired from Kenya Agricultural Research Institute (KARI)-Njoro. Each cultivar was planted in pots (25 seeds pot-1) in forest soil-farm yard manure medium (2:1 v/v). To ensure that flowering dates for all the varieties were synchronized, the late maturing varieties were planted earlier than the early maturing varieties. The plants were maintained outside the green house until flowering (GS61, Zadoks et al., 1974) to simulate field conditions. The plants were fertilized after germination at GS10 (NPK 20:20:0), at tillering GS22 (NPK 20:20:0) and at booting GS41 (urea 46% N) at the rate of 5 g pot-1. Leaf rust (Puccinia recondite) was controlled by spraying with triadimefon (Bayleton®) at tillering (GS 22) at the rate of 1 g L-1 while leaf chewing insects and aphids were controlled with dimethoate (Danadin® 40 EC) at the rate of 1 mL-1. The experiment was repeated over two greenhouse cropping cycles.
Inoculum production and inoculation: Fusarium graminearum was isolated from infected wheat seeds obtained from farmers fields during the 2004 cropping season. The seeds were surface sterilized for 3 min in 5% sodium hypochlorite containing four drops of 20 for 3 min then rinsed three times with distilled water. The kernels were plated on low strength Potato Dextrose Agar (PDA) media amended with mineral salts (Muthomi, 2001). The Fusarium isolates were identified to species level based on synoptic keys by Nelson et al. (1983). Pathogenicity of the different isolates was tested by inculation onto ears of a susceptible wheat variety (Mbuni) according to Muthomi et al. (2002b). Three most pathogenic isolates of Fusarium graminearum were selected for the study. Each isolate was cultured separately on potato dextrose agar and inoculum was multiplied in mung bean liquor medium (Bai and Shaner, 1996). Spore suspension for each isolate was prepared by filtering the aqueous culture through two layers of cheesecloth and spore concentration adjusted to 5x105 spores mL-1. The spore suspensions of the 3 isolates were then mixed to make a composite inoculum and a few drops of 20 added to ensure uniform spore dispersion. Inoculation was done at 50% flowering (GS65) by spraying the ears with the spore suspension using a hand sprayer. Re-inoculation was done after 4 days. Control pots were sprayed with distilled water. The inoculated ears were immediately covered with polythene bags for 48 h to ensure high humidity for infection. Four pots were inoculated per cultivar and the experiment arranged in a complete randomized block design. After inoculation, the plants were allowed to mature inside the greenhouse (22±5°C) to ensure uniform environmental conditions and to avoid damage from birds.
Head blight assessment and effect on grain yield: Head blight severity was assessed visually as the proportion of bleached spikelets based on a 1-9 scale (Miedaner and Perkowski, 1996), where 1= no symptoms, 2 = <5%, 3 = 5-15%, 4 = 16-25%, 5 = 26- 45%, 6 = 46-65%, 7 = 66-85%, 8 = 86-95% and 9 = 96-100% of bleached spikelets. Ten average sized ears per pot were tagged and assessed for disease every 5th day starting with onset of symptoms until yellow ripening (GS87). The mean disease severity value for each replicate pot was calculated by averaging the disease severity values for the different assessment times. The Area Under Disease Progress Curve (AUDPC) was calculated as follows (Shaner and Finney, 1977):
Where, Yi is the disease severity at the ith observation, n is the total number of observations, ti is the time (days) at the ith observation. At maturity, the tagged ears were harvested separately for the different cultivars, dried and threshed by hand for grain weight determination. Seed grains of each cultivar were weighed separately (ten ears per pot). Data was subjected to analysis of variance (ANOVA) using Genstat® for Windows, 6th edition. Where treatment effect was significant, pair-wise treatment mean differences were determined by Tukey Least Significant Difference (LSD) test at 95% confidence limit.
RESULTS
The most widely grown wheat cultivars in the two districts surveyed during the 2004-growing season were Kwale, Mbuni, Chiriku, Nyangumi and Pasa (Table 1). The 8 cultivars challenged with Fusarium head blight by inoculation with F. graminearum significantly differed (p≤0.05) in both disease severity and area under disease progress curve (Table 2).
| Table 1: | Percentage of farmers growing different wheat cultivars in Nakuru and Nyandarua districts of Kenya during the 2004 cropping season |
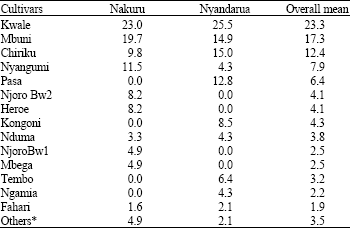 | |
| * Mwamba, Chozi, Popo, Royal | |
| Table 2: | Mean disease severity, 10 ear seed weight (g), 100 seed weight (g) and kernel weight reduction for the 8 wheat cultivars inoculated with Fusarium graminearum |
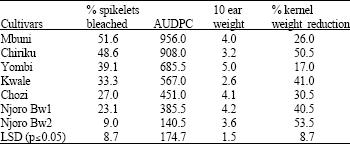 | |
| AUDPC = Area Under Disease Progress Curve | |
The mean disease severity ranged from 9.0% for the least susceptible (Njoro Bw2) to 51.6% for the most susceptible cultivar (Mbuni). The area under disease progress curve varied from 140.5 to 956 for the least and most susceptible cultivars, respectively. Disease severity was highly significantly (p≤0.05) correlated to the area under the disease progress curve (r = 0.85 to 0.99). The cultivars also significantly differed (p≤0.05) in grain weight. The most susceptible cultivars were Mbuni, Chiriku, Yombi, Kwale and Chozi while the least susceptible were Njoro Bw2, Heroe and Njoro Bw1 (Fig. 1). Based on head blight severity and the area under disease progress curve, the 8 wheat cultivars could be grouped into three susceptibility groups (Fig. 1): susceptible (Mbuni and Chiriku), moderately tolerant (Yombi, Kwale and Chozi) and tolerant (Njoro Bw1, Heroe and Njoro Bw2). In the most susceptible cultivars, symptoms appeared as early as 5 days after inoculation and disease development was faster compared to 10 days incubation period for the least susceptible cultivars (Table 3). The most widely grown cultivars in Nakuru and Nyandarua districts, Kenya, were among the highly susceptible and moderately susceptible.
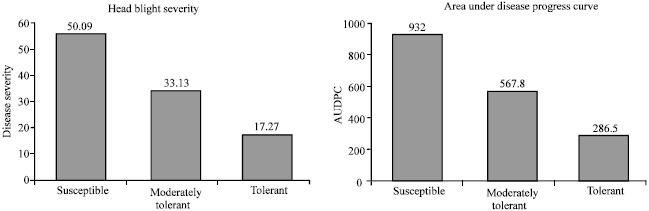 | |
| Fig. 1: | Susceptibility groups of wheat cultivars based on the mean head blight severity and the area under disease progress curve (Susceptible-Mbuni and Chiriku; Moderately tolerant-Yombi, Kwale and Chozi; Tolearant- Njoro Bw1, Heroe and Njoro Bw2) |
| Table 3: | Disease progress on 8 different wheat cultivars inoculated with Fusarium graminearum |
 | |
| Means followed by same letter(s) within columns are not significantly different (p = 0.05) | |
DISCUSSION
The results indicated that the widely grown wheat cultivars in Nakuru and Nyandarua districts of Kenya were susceptible to Fusarium head blight. This suggests that head blight could pose a major threat to wheat production in the two districts in case an epidemic broke out under favourable weather conditions. All the 8 wheat cultivars that were inoculated with Fusarium graminearum were found to be susceptible to head blight. However, the cultivars differed in susceptibility as indicated by the incubation period, disease severity and kernel weight reduction. The results were consistent with findings by Muthomi et al. (2002a) who tested 15 different Kenyan wheat cultivars and found that they were all susceptible to Fusarium head blight. The cultivars could be divided into three susceptibility groups: highly susceptible (Mbuni and Chiriku), moderately susceptible (Yombi, Heroe, Chozi, Kwale and Njoro Bw1) and moderately resistant (Njoro Bw2). The differences in susceptibility among the cultivars could probably be due to inherent genetic resistance factors.
Two types of resistance to FHB of wheat have been reported; resistance to primary infection and resistance to spread of the disease within a spike (Lin et al., 2006; Ma et al., 2006; Wang and miller, 1988; Schroeder and Christensen, 1963). A third mechanism, resistance to deoxynivalenol accumulation has been proposed (Ma et al., 2006; Miller et al., 1985) and deoxynivalenol may play a role in Fusarium head blight pathogenesis (Snijders and Krechting, 1992). Fusarium head blight resistance in wheat has been reported to be a quantitative trait that is controlled by a polygenic system. Effects of dominance of genes probably influence FHB resistance, but additive effects appear to be important and resistance genes can be accumulated (Shen and Ohm, 2006; Bai et al., 1999). Chromosomal location of FHB resistance genes/loci in the wheat genome has been mapped using RFLP and AFLP methods and recombinant inbred lines. Quantitative trait loci of resistance to FHB have been preliminarily mapped on the following chromosomes: 1B, 2A; 2AL, 2B, 3 BS, 3A, 3B, 3D, 4A, 4B, 5A, 5B, 5D, 6A, 6B and 6D (Lin et al., 2006; Ma et al., 2006; Shen and Ohm, 2006; Mardi et al., 2005; Shen et al., 2003). Growing of wheat cultivars resistant to Fusarium sp. is the most economic, environment-friendly and effective method of disease control. Sources of resistance have been found in China, South America and Czech Republic (Lin et al., 2006; Mersterhazy et al., 1999; Mersterhazy, 1995). Currently, there are no wheat varieties with high level of resistance to FHB although some varieties have useable levels of partial resistance that limit yield loss and mycotoxins accumulation (Shen and Ohml, 2006; Pereyra and Dill-Macky, 2004). Cultivar Sumai 3 from China is reported to be the best source of resistance (Kolb et al., 2001) and its quantitative trait loci located on the distal end of the short arm of chromosome 3B (Liu et al., 2005) is stably expressed in different environments and in different genetic backgrounds (Shen et al., 2003; Zhou et al., 2002). It combines good resistance to both type I and type II infection and gives one of the lowest levels of DON in the kernels following infection. Good resistance to FHB can also be found in wild relatives of wheat and related triticale species. A recent study by Shen and Ohm (2006) reported that FHB resistance on chromosome 7E of wheatgrass Lonhopyrum elongatum confers a large effect and has an additive effect with chromosome 3B of chinese wheat cultivar Sumai 3. Such incorporation of FHB resistance from related species is critical to improving the level of resistance, enhancing genetic diversity and providing continuing resistance.
This study could be useful in providing information to wheat producers in wheat growing areas of Kenya on selection of cultivar during planting. This is critical considering that most of the cultivars grown in Nakuru and Nyandarua districts (as well as other wheat producing regions in Kenya) are susceptible to FHB. Therefore the selected cultivar will among other factors determine disease infection levels and possible mycotoxin contamination besides the economic implications of FHB management or resultant yield loss. The study also highlights the need for continued breeding for resistance against FHB causing fusaria. Further studies are needed to determine the type of resistance in local wheat cultivars and to determine whether the genes involved could be useful in development of new cultivars.
ACKNOWLEDGMENT
This research was funded by the International Foundation for Science (IFS) grant No.E/3654-1.
REFERENCES
- Bai, G.H. and G. Shanner, 1996. Variation in Fusarium graminearum and cultivar resistance to wheat scab. Plant Dis., 80: 975-979.
CrossRefDirect Link - Nganje, W.E., D.A. Bangsund, F.L. Leistritz, W.W. Wilson and N.M. Tiapo, 2004. Regional economic impacts of Fusarium head blight in wheat and barley. The Rev. Agric. Eco., 26: 332-347.
Direct Link - Bai, G., F.L. Kolb, G. Shaner and L.L. Domer, 1999. Amplified fragment length polymorphism markers linked to a major quantitave trait locus controlling scab resistance in wheat. Phytopathology, 89: 343-348.
Direct Link - Bateman, G.L., G. Murray, R.J. Gutteridge and H. Coslkun, 1998. Effects of method of straw disposal and depth of cultivation on populations of Fusarium spp. in soil and on brown foot rot in continuous winter wheat. Ann. Applied Biol., 132: 35-47.
CrossRefDirect Link - Bateman, G.L. and G. Murray, 2001. Seasonal variations in populations of Fusarium species in wheat-field soil. Applied Soil Ecol., 18: 117-128.
Direct Link - Bateman, G.L., 2005. The contribution of ground-level inoculum of Fusarium culmorum to ear blight of winter wheat. Plant Pathol., 54: 299-307.
Direct Link - Brennan, J.M., D. Egan, B.M. Cooke and F.M. Doohan, 2005. Effect of temperature on head blight of wheat caused by Fusarium culmorum and F. graminearum. Plant Pathol., 54: 156-160.
CrossRefDirect Link - Champeil, A, J.F. Fourbet, T. Dore and L. Rossignol, 2004. Influence of cropping system on Fusarium head blight and mycotoxin levels in winter wheat. Crop Prot., 23: 531-537.
CrossRefDirect Link - D'Mello, J.P.F., C.M. Placinta and A.M.C. MacDonald, 1999. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol., 80: 183-205.
CrossRefDirect Link - Doohan, F.M., J. Brennan and B.M. Cooke, 2003. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol., 109: 755-768.
Direct Link - Fernando, W.G.D., T.C. Paulitz, W.L. Seaman, P. Dutilleul and J.D. Miller, 1997. Head blight gradients caused by Gibberella zeae from area sources of inoculum in wheat field plots. Phytopathology, 87: 414-421.
Direct Link - Goswami, R.S. and H.C. Kistler, 2004. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol., 5: 515-525.
CrossRefDirect Link - Heier, T., S.K. Jain, K.H. Kogel and J. Pons-Kühnemann, 2005. Influence of N-fertilization and fungicide strategies on Fusarium head blight severity and mycotoxins content in winter wheat. J. Phytopathol., 153: 551-557.
Direct Link - Jones, R.K., 2000. Assessments of Fusarium head blight of wheat and barley in response to fungicide treatment. Plant Dis., 84: 1021-1030.
Direct Link - Kolb, F.L., G.H. Bai, G.J. Muehlbauer, J.A. Anderson, K.P. Smith and G. Fedak, 2001. Symposium on genetic solutions to Fusarium head blight in wheat and barley: Challenges, opportunities and imperatives. Host plant resistance genes for Fusarium head blight: Mapping and manipulation with molecular markers. Crop Sci., 41: 611-620.
- Liu, S., M. Zhang, M.O. Pumpphrey, R.W. Stack, B.S. Gill and J.A. Anderson, 2005. Complex microcolinearity among whaet, rice and barley revealed by fine mapping of genomic region harbouring a major QTL for resistance to Fusarium head blight in wheat. Func. Intergrative Genomics, 6: 83-89.
CrossRef - Llorens A., R. Mateo, M. Hinojo, F.M. Valle-Algarra, M. Jimenez, 2004. Influence of environmental factors on the biosynthesis of type B trichothecenes by isolates of Fusarium sp. from Spanish crops. Int. J. Food Microbiol., 94: 43-54.
CrossRefDirect Link - Ma, H.X., K.M. Zhang, L. Gao, H.G. Chen, Z.X. Cai and W.Z. Lu, 2006. QTL for the resistance to wheat Fusarium head blight and deoxynivalenol accumulation in Wangshuibai under field conditions. Plant Pathol., 55: 739-745.
CrossRef - Magan, N., R. Hope, A. Colleate and E.S. Baxter, 2002. Relationship between growth and mycotoxin production by Fusarium species, biocides and environment. Eur. J. Plant Pathol., 108: 685-690.
Direct Link - Mardi, M., H. Buerstmayr, B. Ghareyaziel, M. Lemmens, S.A. Mohammadi, R. Nolz and P. Ruckenbauer, 2005. QTL analysis of resistance to Fusarium head blight in wheat using a `Wangshuibai-derived population. Plant Breed., 124: 329-333.
Direct Link - Mentewab, A., H.N. Rezanoor, N. Gosman, A.J. Worland and P. Nicholson, 2000. Chromosomal location of Fusarium head blight resistance genes and analysis of the relationship between resistance to head blight and brown foot rot. Plant Breed., 119: 15-20.
CrossRefDirect Link - Mesterhazy, A., 1995. Types and components of resistance to Fusarium head blight of wheat. Plant Breed., 114: 377-386.
CrossRefDirect Link - Mesterhazy, A., T. Bartok, C.G. Miropcha and R. Komoroczy, 1999. Nature of wheat resistance to Fusarium head blight and the role of deoxynivalenol for breed. Plant Breed., 118: 97-110.
CrossRefDirect Link - Miedaner, T. and J. Perkowski, 1996. Correlation among Fusarium culmorum head blight resistance, fungal colonization and mycotoxin content in winter rye. Plant Breed., 115: 347-351.
Direct Link - Miedaner, T., 1997. Breeding wheat and rye for resistance to Fusarium disease. Plant Breed., 116: 201-220.
Direct Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. 1st Edn., Pennsylvania State University Press, University Park, University Park, PA., USA., ISBN-13: 978-0271003498, Pages: 226.
Direct Link - Mishraa, P.K., R.T. Foxa and A. Culhama, 2002. Restriction analysis of PCR amplified nrDNA regions revealed intraspecific variation within populations of Fusarium culmorum. Microbiol. Lett., 215: 291-296.
Direct Link - Muthomi, J.W., E.C. Oerke, H.W. Dehne and E.W. Mutitu, 2002. Susceptibility of Kenyan wheat varieties to head blight, fungal invasion and deoxynivalenol accumulation inoculated with Fusarium graminearum. J. Phytopathol., 150: 30-36.
CrossRefDirect Link - Muthomi, J.W., E.C. Oerke, E.W. Mutitu, A. Schade-Schutze and H.W. Dehne, 2002. Variation among Fusarium species and isolates infecting wheat ears based on aggressiveness, mycotoxin production and RAPD-PCR analysis. J. Plant Dis. Prot., 109: 462-477.
Direct Link - Parry, D.W., P. Jenkinson and L. McLeod, 1995. Fusarium ear blight (scab) in small grain cereals. Plant Pathol., 44: 207-238.
Direct Link - Pereyra, S.A., R. Dill-Macky and A.L. Sims, 2004. Survival and inoculum production of Gibberella zeae in wheat residue. Plant Dis., 88: 724-730.
CrossRefDirect Link - Ramirez, L., S. Chulze and N. Magan, 2004. Impact of environmental factors and fungicides on growth and deoxinivalenol production by Fusarium graminearum isolates from Argentinian wheat. Crop Protect., 23: 117-125.
CrossRefDirect Link - Shaner, G. and R.E. Finney, 1977. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in knox wheat. Phytopathology, 67: 1051-1056.
CrossRefDirect Link - Shen, X., M. Zhou, W. Lu and H. Ohm, 2003. Detection of Fusarium head blight resistance QTL in a wheat population using bulk segregant anal. Theor. Applied Genet., 106: 1041-1047.
Direct Link - Windels, C.E., 2000. Economic and social impacts of Fusarium head blight: Changing farms and rural communities in the northern Great Plains. Phytopathology, 90: 17-21.
PubMed - Wisniewska, H. and K. Kowalczyk, 2005. Resitance of cultivars and breeding lines of spring wheat to Fusarium culmorum and powdery mildew. J. Applied Genet., 46: 35-40.
PubMedDirect Link - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link - Zhou, W.C., F.L. Kolb, G. Bai, G. Shaner and L.L. Domier, 2002. Genetic analysis of scab resistance QTL in wheat with microsatellite and AFLP markers. Genome, 45: 719-727.
PubMedDirect Link