
A.G.D. Delima
Department of Biological Sciences and Environmental Studies, College of Science and Mathematics, University of the Philippines Mindanao, Mintal, Davao City, Philippines
O.P. Damasco
Laboratory of National Plant Genetic Resources, Institute of Plant Breeding, University of the Philippines Los Ba�os, Laguna, Philippines
A.U. Novero
Department of Biological Sciences and Environmental Studies, College of Science and Mathematics, University of the Philippines Mindanao, Mintal, Davao City, Philippines
Asian Journal of Plant Sciences
Year: 2015 | Volume: 14 | Issue: 2 | Page No.: 66-71







ABSTRACT
The value of sago palm (Metroxylon sagu Rottb.) as a high starch yielding crop has not been fully realized yet but at risk to human-mediated and environmental threats that necessitate its conservation. Attempts were made to investigate cryopreservation as an option for long-term storage of sago palm zygotic embryos using simple desiccation technique. Zygotic embryos were aseptically removed from white sago palm seeds. Laminar desiccation (1-3 h) and silica gel drying (1-7 h for 50, 100 and 150 g) were employed to reduce moisture content without losing much viability. Viability was assessed through tetrazolium test and germination in modified Murashige and Skoog media. Desiccation of embryos in 100 g silica gel for 6 h was the selected pre-treatment to cryopreservation. The critical moisture content of embryos was at 21 with 55% viability. With storage in liquid N, staining showed 14-46.7% partial viability after 2 and 24 h, respectively. High desiccation sensitivity of immature sago embryos hindered its direct storage to liquid nitrogen which suggests complementing simple dehydration techniques with cryoprotectants. Cryopreservation of sago palm remains a potential and progressing approach for future long-term conservation and management of this versatile starch crop.
PDF Abstract XML References Citation
Received: June 23, 2015;
Accepted: October 15, 2015;
Published: October 27, 2015
How to cite this article
A.G.D. Delima, O.P. Damasco and A.U. Novero, 2015. Utilization of Sago Palm (Metroxylon sagu Rottb.) Zygotic Embryos for Cryopreservation. Asian Journal of Plant Sciences, 14: 66-71.
DOI: 10.3923/ajps.2015.66.71
URL: https://scialert.net/abstract/?doi=ajps.2015.66.71
DOI: 10.3923/ajps.2015.66.71
URL: https://scialert.net/abstract/?doi=ajps.2015.66.71
INTRODUCTION
Sago palm (Metroxylon sagu Rottb.), one of the highest starch yielding crops available is less cultivated in the Southern region of the Philippines than other countries of Southeast Asia (Quat Ng, 2007). It is usually associated with lumads or indigenous people (Flores, 2009). The long life span, late maturity and reproductive method of sago also contribute to genetic vulnerability (Rounds, 2007). Sago serves as staple food for some local people. In Agusan province, cutting down of sago is rampant to give way to quickly-maturing cash crops and converted to rice lands (Flores, 2008).
In view of conservation, sago palm’s recalcitrant seeds cannot be dried and stored in cold rooms. Field gene banking offer a means of preservation of the natural stands (e.g. sago stands in Agusan Marsh Sanctuary as a protected area). However, this method exposes sago palms to natural disasters like typhoons and also made them prone to pests and diseases which may lead to losses of genotypes (Mroginski et al., 2008). Aside from that, a field gene bank requires large space, high labor and financial costs too (Kim et al., 2006; Bandupriya et al., 2010).
The drawback of field gene banking can be complemented with an alternative strategy known as cryopreservation for long-term conservation, applicable to recalcitrant where seeds for ex situ storage are limited (Bunn et al., 2007). Cryopreservation is an important alternative and relatively inexpensive method that complements in situ strategies (Efendi and Litz, 2003). It is useful among members of palm family such as oil palm and coconut (Panis and Lambardi, 2005). This study is the first reported attempt on sago palm cryopreservation for long-term conservation, specifically: to determine the effective method of reducing moisture content using simple desiccation pre-treatment of zygotic embryos and to evaluate the effects of reduced moisture content on the viability and germination of embryos after cryopreservation.
MATERIALS AND METHODS
Collection of sago palm seeds: Sago palm fruits were collected in La Filipina, Tagum City, Davao del Norte, Mindanao, Philippines. Only white seeds were available and were obtained. Then, series of surface sterilization steps were carried out prior to aseptic removal of zygotic embryos.
Desiccation pre-treatment: Prior to cryopreservation, the moisture content of 4-5 mm zygotic embryos was reduced through simple desiccation techniques by using silica gel and laminar flow hood drying. Fresh weight was recorded before oven-drying for 48 h in 102°C and then the dry weight of the embryos were obtained. This served as control. The duration (h) of desiccation appropriate to reduce the moisture content of the embryos was tested for both pre-treatments. In silica gel drying, the embryos were dried for 1, 2, 3, 4, 5 and 6 h with three amounts of silica gel used: 50, 100 and 150 g for each time of desiccation. Thus, there were 18 treatments for silica gel drying. For the second method, the zygotic embryos were exposed in the stream of air in the laminar flow hood with airflow velocity average of 0.3-0.5 m sec–1 for 1-3 h only.
Tetrazolium test: Viability of embryos after desiccation was examined using 1% triphenyl tetrazolium chloride (TTZ) (Sugimura and Murakami, 1990) for 12 h under dark conditions (Kumaunang, 2002). The embryos that turned red were considered viable while non-viable if tissues were abnormally stained or not stained at all (Patil and Dadlani, 2009). The percentage of viable embryos and the average per treatment was determined after staining. From the results of the viability test, the desiccation treatment with the highest percentage viability and low moisture content was selected for cryopreservation.
Cryopreservation of sago palm: After extraction and sterilization of sago embryos, 10 embryos per replicate were stored in six sterile 50 mL Falcon tubes. The cryocanes were submerged in the cryotank for minimum of 2 h and maximum of 24 h. After 2 and 24 h of storage in liquid nitrogen (-196°C), cryotubes were pulled out and thawed rapidly in hot water bath at 40°C for 2 min. Then, the tubes were stabilized in room temperature for 5 min. The tubes were then transferred to the laminar flow hood for inoculation of cryo-preserved embryos. Modified Murashige and Skoog (MS) medium supplemented with 50 μM BAP and 100 μM NAA was used. On the other hand, embryonic axes were directly inoculated without storage in liquid nitrogen. Viability and moisture content of two set-ups were compared. Simple desiccation techniques are important tools that should initially be investigated prior to use of cryoprotectants. Cyroprotectants include dimethylsulfoxide (DMSO), glycerol and ethylene glycol which can increase the recovery of the cryo-preserved zygotic embryos (Normah and Makeen, 2008).
Statistical analysis: The experiments were conducted in a Completely Randomized Design (CRD) with six trials, 3 replicate each and 10 embryos per replicate. The data was subjected to Univariate Analysis of Variance. Then, for multiple comparisons of the means, Least Significant Difference (LSD) was applied (Gomez and Gomez, 1984).
RESULTS
Effect of laminar flowhood desiccation on MC and viability of embryos: The percentage of Moisture Content (MC) and viability of sago embryos differed with the control (Table 1). Significant reductions in moisture content from 92.9 (control) to 89 and 71% after 1 and 3 h exposures in laminar flow hood, respectively were observed. With further drying of embryos, moisture content declined from 70% after 3 h and 30% after 4 h. After 5 h, 28.6% MC was achieved and went down to 16.7% MC after 6 h. Viability of the embryos was greatly reduced even after a 3 h desiccation period. Greater loss of viability was observed with longer desiccation time.
Effect of silica gel desiccation on percentage of MC and viability of embryos: The percentage of Moisture Content was reduced on the average from 92-16.7%, being the lowest moisture content after 7 h dehydration in 150 g silica gel. Increasing duration of drying in silica gel from 1-7 h resulted to corresponding reductions in moisture content of the embryos. Moisture content of the embryos also decreased with the increasing amount of silica gel (Table 2).
In non-desiccated embryos (control), viability decreased from 100-30% (7 h in 50 g silica gel). In terms of viability, severe loss of viability of embryos was found in laminar flow hood (20%) desiccation than in silica gel drying (60-80%). Though faster loss of water content was observed in silica gel, its effect on the loss of viability of embryos was not drastic and rapid due to a more uniform and controlled drying unlike the laminar flow hood. Between the effect of laminar flow hood drying and silica gel dehydration, sago palm embryos had higher tolerance under silica gel even at longer duration of exposure up to 7 h compared with the laminar flow hood exposure up for 3 h.
| Table 1: | Effect of duration of laminar flowhood desiccation on percentage of MC and viability of sago embryos |
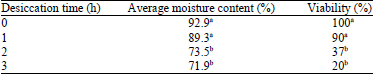 | |
| MC: Moisture content, Means with different letters are significantly different from each other at 0.05 level | |
| Table 2: | Moisture content and viability of embryos desiccated in varying amounts of silica gel at certain time in hours |
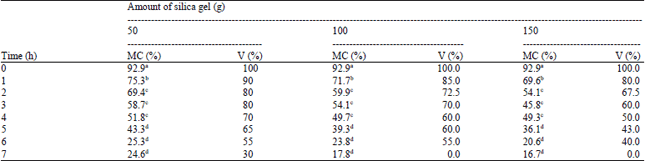 | |
| MC: Moisture content, V: Viability, Means with different letters are significantly different from each other at 0.05 level | |
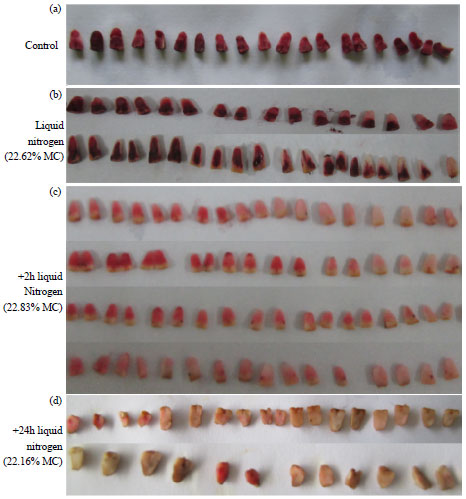 | |
| Fig. 1(a-d): | Viability of sago palm embryos with or without liquid nitrogen compared with control, Viability assessment by tetrazolium staining, (a) Viable-completely stained red embryos, (b) Partial viability-only plumule or radicle area are stained red; patches of stains only, light red staining over the embryos, (c) Dead-very light pink superficial staining and (d) Completely unstained |
Results revealed that 6 h of drying in 100 g of silica gel seemed optimal in lowering the moisture content to 23.8%, with viability of 55%, the critical moisture level for sago embryos with almost 50% reduction in viability.
Viability test: The general staining patterns of the embryos varied and decreased in intensity in redness as the length of time of desiccation increased. The formation of red formazan varied from whole embryos to patches and spots or faint surface stains in 1-3 h laminar drying and 1-7 h in various silica gel exposures. Cryopreservation for 2 and 24 h (Fig. 1) showed partial viability of the embryos which indicated low tolerance of desiccated embryos after storage in liquid nitrogen with faint red or very light pink color being noted frequently.
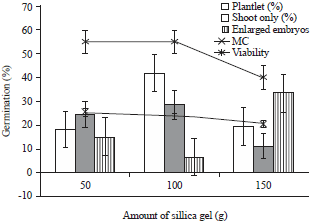 | |
| Fig. 2: | Effect of percentage of MC on viability and germination of embryos after 6h exposure in different amount of silica gel |
Germination test: Direct germination of embryos after 6 h drying resulted to reduced complete germination. Highest germination of embryos into plantlet (41%) as well as shoot formation (21%) was obtained in 100 g silica gel drying. The remaining cultures consisted of enlarged embryos. Further, lower germination was observed in 50 and 150 g silica gel drying. Viability was higher than germination percentage (Fig. 2). After 6 h silica gel drying at different amounts, shoots were noted to be shorter than normal and some embryos enlarged, while others remained to be whitish in the medium.
Effect of cryopreservation of zygotic embryos: Sago embryos, 4-5 mm in length were desiccated in 100 g silica gel for 6 h prior to cryopreservation. This proved to be the effective method in reducing the moisture content to its lowest level (21%). However, only 40% viability was retained and 24% of the embryos germinated after in vitro germination test. Storage in liquid nitrogen reduced the viability of embryos from 55-47 and 14% after 2 and 24 h, respectively. Faint red or very light pink colors were noted more frequently. The radicle portions near the base of the embryos were observed unstained compared to control which was 100% stained. Meanwhile, the 6h drying in silica gel as pre-treatment yield partial germination with 31.6% shoot formation and 26.3% plantlet regeneration (Fig. 3). Few cultures were observed to have developed roots alone. About 24% of the remaining cultures became enlarged.
DISCUSSION
The relationship between viability and moisture content over desiccation time was directly proportional. Reduced moisture in laminar flow hood caused drastic loss on viability of the embryos.
In laminar flow hood dehydration, the loss of moisture content was gradual in contrast to silica gel desiccation. The varied rate of drying between the two techniques may be influenced by the relative humidity of the environment (Dumet et al., 1993), as such with high air humidity the dehydration was comparably slower and not precise drying in air for small embryos (Assy-Bah and Engelmann, 1992). Unlike silica gel drying, embryos were contained in a sealed container, thereby not exposing the embryos to relative humidity which could affect reduction of moisture content. In addition, silica gel provided air-tight close condition which led to slower reduction in viability. Viability ranged from 50-70% after 4 h and 43-65% after 5 h. Then, after 6 h, viability was further reduced to 40-55%.
Drying time in silica gel was longer than laminar flowhood drying. This technique of dehydration allowed sago palm embryos to adapt to water stress and had a higher tolerance. In oil palm, silica gel was the favoured drying method over laminar flowhood. Slower rate of drying may have caused tolerance to desiccation over time (Kim et al., 2006).
The levels of moisture content after desiccation at 23% can be classified under type III water, membrane-bound type of water (Vertucci and Farrant, 1995). The removal of bound water results to mechanical damage on seeds (Kumaunang, 2002) affecting the viability of the zygotic embryos.
The critical period of drying was 6 h in 100 g of silica gel because viability was greatly reduced beyond this. Reduced recovery, only about 55% of embryos were darkly stained, indicated that prolonged exposure of embryos to silica gel for 6 h caused the loss of germination capability of embryos. Reduction in water level can cause structural changes not just at tissue level but even cellular or subcellular level (Kumaunang, 2002). Further, abnormal germination was observed such as enlarged embryos, radicle or shoot development alone. Similar abnormal growth was observed in Nothapodytes nimmoniana with only radicle (Radha et al., 2009).
Staining patterns indicate the activity of living cells such as respiration of mitochondria (Normah and Makeen, 2008). Stains were no longer dark red, which indicated that the embryos’ viability and enzymatic activity may have been severely affected after cryopreservation (Kumaunang, 2002). The reduced moisture content of embryos to 23% was possibly not the optimum moisture content for storage in liquid nitrogen. In cryogenic storage of herbaceous peony (Kim et al., 2006), at 26% moisture level after 1 h desiccation by air drying, 66% post-thaw re-growth was observed. The optimum moisture content ranged from 8-20% but varied depending on species (Normah and Makeen, 2008). This indicated that not all sago palm embryos were able to tolerate freezing in liquid nitrogen after 2 h of storage. Moisture content was also still high which may have not succeeded in preventing the formation of intracellular ice crystals leading to the death of more than half of the cryo-preserved embryos.
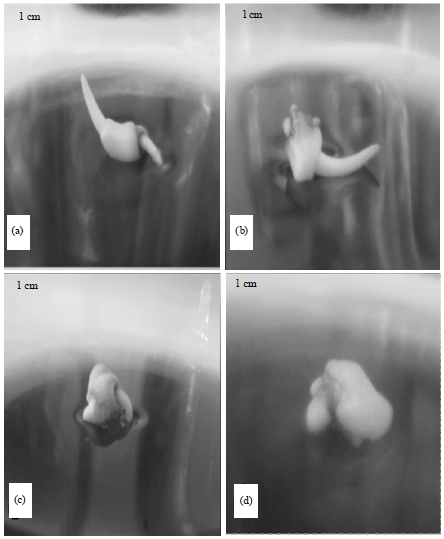 | |
| Fig. 3(a-d): | Direct recovery of sago palm embryos after drying in 100 g silica gel for 6 h (a) Plantlet, (b) Shoot only, (c) Root emergence and (d) Enlarged embryos |
The present study established that 23% was the critical moisture content of sago palm embryos. Initial results showed that embryos suffered from cryo-exposure and were not able to germinate despite the viability shown after tetrazolium test. Thus, the desiccation technique did not work for sago embryos. Further reduction on the moisture content with the aid of cryoprotectants such as dimethyl sulfoxide (DMSO) or other high osmoticum via vitrification are suggested for further study. These techniques may well be useful to increase the tolerance of embryos from desiccation and cryogenic stresses. Vitrification is potentially applicable as it worked on embryos and axes with high desiccation sensitivity like Citrus microcarpa (Normah and Makeen, 2008).
It is also important to consider the physiological maturity of the embryos. Sago palm’s seed maturity and characteristics are not extensively studied yet. The embryos sourced from white seeds were not mature enough compared with black seeds. Zygotic embryos are heterozygous in cross-pollinating plants as in the case of sago palms (Kim et al., 2006). The importance of right developmental stages of embryos is critical for the success of cryostorage (Normah and Makeen, 2008). White seeds are considered to be immature (Jong, 1995), optimization is needed to increase plant regeneration capacity. Cryostorage of mature black seeds is suggested as the histological aspect certainly affects resistance to dehydration (Dumet et al., 1993). Further, the protocol for sago palm may need to be reviewed such as using alternative for the tubes used in storing the embryos for cryopreservation. In rubber tree, there was improved survival rate when embryos were stored using the aluminum envelopes instead of cryotubes (Normah and Makeen, 2008).
Zygotic embryos of white sago palm seeds are found desiccation-sensitive, wherein 21% moisture content is critical losing almost half of the total viability. Being desiccation-sensitive, simple desiccation techniques does not suit the purpose of pre-treating embryos prior to cryopreservation of zygotic embryos of sago palm. Tolerance to desiccation and cryo-exposure may be improved with the use of cryoprotectants. Sago palm being a recalcitrant species is difficult to optimize for ex situ techniques such as cryopreservation.
This study recommends further investigation on the response of zygotic embryos to other pre-treatment techniques for the cryopreservation of sago palm zygotic embryos.
ACKNOWLEDGMENTS
This study was a portion of the MS thesis of A.G. Delima funded by DOST-ASTHRDP-SEI. The authors also acknowledge UPLB Graduate School and DOST-UP Mindanao through Sago Project 2.5. The provisions of Prof. Teresita H. Borromeo and Dr. Pompe Sta. Cruz are greatly appreciated.
REFERENCES
- Assy-Bah, B. and F. Engelmann, 1992. Cryopreservation of immature embryos of coconut (Cocos nucifera L.). Cryo Lett., 13: 67-74.
Direct Link - Bandupriya, H.D.D., S.C. Fernando, J.L. Verdeil and B. Malaurie, 2010. Cryopreservation of encapsulated plumules of coconut: Effect of transport/store conditions. Asia-Pac. J. Mol. Biol. Biotechnol., 18: 135-137.
Direct Link - Bunn, E., S. Turner, M. Panaia and K.W. Dixon, 2007. The contribution of in vitro technology and cryogenic storage to conservation of indigenous plants. Aust. J. Bot., 55: 345-355.
CrossRefDirect Link - Dumet, D., F. Engelmann, N. Chabrillange and Y. Duval, 1993. Cryopreservation of oil palm (Elaeis guineensis Jacq.) somatic embryos involving a desiccation step. Plant Cell Rep., 12: 352-355.
CrossRefDirect Link - Flores, D.M., 2008. The versatile sago (Metroxylon sagu Rottb.) and its green potential for Mindanao. Banwa, 5: 8-17.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons, New York, USA., ISBN-13: 9780471870920, Pages: 680.
Direct Link - Kim, H.M., J.H. Shin and J.K. Sohn, 2006. Cryopreservation of somatic embryos of the herbaceous peony (Paeonia lactiflora Pall.) by air drying. Cryobiology, 53: 69-74.
CrossRefPubMedDirect Link - Mroginski, L.A., P.A. Sansberro, A.M. Scocchi, C. Luna and H.Y. Rey, 2008. A cryopreservation protocol for immature zygotic embryos of species of Ilex (Aquifoliaceae). Biocell, 32: 33-39.
Direct Link - Sugimura, Y. and T. Murakami, 1990. Structure and function of the haustorium in germinating coconut palm seed. Jpn. Agric. Res. Quart., 24: 1-14.
Direct Link