
S.O. Afuape
National Root Crops Research Institute, Umudike, P.M.B. 7006, Umuahia, Abia State, Nigeria
I.I.M. Nwankwo
National Root Crops Research Institute, Umudike, P.M.B. 7006, Umuahia, Abia State, Nigeria
R.A. Omodamiro
National Root Crops Research Institute, Umudike, P.M.B. 7006, Umuahia, Abia State, Nigeria
J.C. Njoku
National Root Crops Research Institute, Umudike, P.M.B. 7006, Umuahia, Abia State, Nigeria
C.L. Ogbonna
National Root Crops Research Institute, Umudike, P.M.B. 7006, Umuahia, Abia State, Nigeria
D.C. Uzuegbu
National Root Crops Research Institute, Umudike, P.M.B. 7006, Umuahia, Abia State, Nigeria
International Journal of Plant Breeding and Genetics
Year: 2015 | Volume: 9 | Issue: 4 | Page No.: 206-217







ABSTRACT
Sweetpotato breeding in Nigeria and many other countries has largely focused on increasing productivity on farmers’ fields and for fresh root consumption. In order to target the breeding program to processing end-use, the variability, location performance, Genotype-by-Environment Interaction (GEI), heritability estimates and relationships among important root processing traits among elite sweetpotato lines were studied in two locations. The genotypes exhibited significant (p<0.05) genotypic variability at each location for dry matter, starch content, flour content and fresh root peel-loss. The combined location analyses also showed that the expression of the traits was under the influence of genotype, environment and GEI. At Makurdi, starch content ranged 14.02-20.53%, while the range of dry matter, flour content and peel-loss were 20.48-37.73, 18.95-31.00 and 11.69-28.40%, respectively. At Umudike, starch was 17.58-23.00%, dry matter was 24.16-34.17%, flour content was 21.34-32.32% and peel-loss was 18.17-24.01%. Correlation study showed that dry matter was moderately correlated to starch (r = 0.53 at Makurdi and r = 0.52 at Umudike) and flour content (r = 0.79 at Makurdi and r = 0.76 at Umudike). Genotypes and traits biplot revealed that NRSP/05/1B, Ex-Igbariam and CIP 440163 largely influenced dry matter, while elevated flour content and peel-loss were related to Ex-Igbariam, CIP 440163 and TIS 87/0087. For heritability study, all the traits showed moderate to high heritability (broadsense) in each location and across locations. This work shows how to identify genotypes with desirable processing traits for release as new varieties. It also enhances the selection of new parent lines for developing new population for processing end-use.
PDF Abstract XML References Citation
Received: June 25, 2015;
Accepted: August 05, 2015;
Published: September 15, 2015
How to cite this article
S.O. Afuape, I.I.M. Nwankwo, R.A. Omodamiro, J.C. Njoku, C.L. Ogbonna and D.C. Uzuegbu, 2015. Targeted Breeding for Sweetpotato-Based Enterprises: Variability, Genotype-by-Environment Interaction, Heritability and Correlation Studies of Important Sweetpotato Root Processing Quality Traits. International Journal of Plant Breeding and Genetics, 9: 206-217.
DOI: 10.3923/ijpbg.2015.206.217
URL: https://scialert.net/abstract/?doi=ijpbg.2015.206.217
DOI: 10.3923/ijpbg.2015.206.217
URL: https://scialert.net/abstract/?doi=ijpbg.2015.206.217
INTRODUCTION
Sweetpotato is a root crop with immense but untapped potentials. It was probably introduced to Nigeria by Spanish and Portuguese explorers in the 18th century (Agbo and Ene, 1994). Its long presence in Nigeria has made it spread and adapted to all agro-ecologies where it is presently cultivated. The short growing period of four months from planting to harvesting and its ability to produce in marginal and waste lands (Fawole, 2007), the ease and lower cost of its field management compared to yam and cassava (Chukwu, 2001), its drought tolerant ability, as well as its high productivity per unit area have endeared sweetpotato to crop farmers. In the last two decades, sweetpotato production has increased rapidly from less than 1.0 million t in the 1990s to over 3.0 million t in 2013 (FAOSTAT., 2013). Today, sweetpotato is produced in all states of the federation with different intensities.
Sweetpotato breeding in Nigeria has largely targeted the development of new varieties with high yielding ability, virus resistance and culinary attributes. This has biased sweetpotato breeding towards enhancing the production of fresh roots for consumption ends. Little research has been directed towards the development of varieties with good processing/industrial attributes. This is evident in the low utilization of the crop for commercial processing. To sustain the increasing trend in sweetpotato production, processing of sweetpotato must be enhanced to utilize produced roots. Important sweetpotato products in Nigeria include sweetpotato fries, non-alcoholic beverage (kunnu), chips, sweetpotato flour and kambar (sliced, parboiled and dried roots). Elsewhere, known sweetpotato products include noodles, bakery foods, snack foods, confectionery products, alcohol production and in the brewing industries (Chen et al., 2003a, b; Dansby and Bovell-Benjamin, 2003). The quality and acceptability of these products are influenced by such processing traits like dry matter, starch content and quality, flour content and peel-loss (for manual processing, which is the dominant micro- and small-scale root and tuber processing method in Nigeria) (Avula, 2005). Therefore, increasing the industrial utilization of sweetpotato can be enhanced by developing varieties with high starch content and quality, high dry matter and high flour content. However, breeding progress depends largely on the availability of wide variation in the breeder’s germplasm, presence and extent of genotype-by-environment interaction on trait expression and the heritability of the traits under selection. Thus, to be able to develop new varieties that are targeted towards adoption for enhanced processing through breeding, there is need to study the variability within the elite germplasm for these traits, understand how the traits respond in different locations, understand the heritability of the traits, as well as study the relationships that exist among the traits. This paper presents the study of these traits in elite sweetpotato breeding lines for rapid selection of promising lines for varietal release.
MATERIALS AND METHODS
Locations: The study was conducted in two locations; Makurdi and Umudike.
Umudike: The Umudike location was at the Experimental Farm of the National Root Crops Research Institute, Umudike, Nigeria within the rainforest belt. Umudike was located on longitude 07°34' E, latitude 05°29' N and altitude 122 m above sea level. The annual rainfall was 2076.8 mm and the general soil type was described as an ultisol.
Makurdi: Makurdi was located within the Southern guinea savannah ecology with total annual rainfall of 1211.48 mm. It was on longitude 08°45' E, latitude 07°45' N and altitude of 98 m above sea level.
Experimental design and layout: Using a randomized block design with three replications, twelve advanced sweetpotato breeding lines in Makurdi and fourteen breeding lines in Umudike were evaluated during the 2013 cropping season for their agronomic and processing attributes. At each site, plot size of 9 m2 (3×3 m) was used, while the plant spacing was 0.3 m along the row and 1m between rows, giving plant density of 33,333 plants ha–1. Fertilizer application (NPK 15:15:15) was carried out at 4 weeks after planting immediately after the first weeding at the rate of 400 kg ha–1. Agronomic data was collected for analyses (data not presented) while medium-sized fresh root samples were collected from each plot for dry matter content (%), starch content (%), flour content (%) and peel-loss (%) determination. The root samples were carefully bagged, labeled and transferred to the laboratory for immediate analyses.
Laboratory procedure: Standard methods of the Official Methods of Analytical Chemist (AOAC., 2010) were used for the determination of the dry matter and starch contents. For flour content, 1 kg fresh root was measured, dried to constant weight, ground to flour and sieved. The flour content was taken as the flour weight percentage of the unpeeled fresh weight. For peel-loss determination, roots from the same genotype from the three replications were bulked and the fresh root weight (kg) measured. The fresh roots were carefully peeled and the weight of the peels per genotype was measured. Peel-loss was calculated as the peel weight percentage of the unpeeled fresh root weight. The analysis for each trait was replicated three times.
Statistical analyses: Few of the advanced sweetpotato breeding lines evaluated for processing attributes were not present in the two locations and the number of genotypes evaluated in each location also differed-twelve in Makurdi and fourteen in Umudike. These disparities necessitated the analysis of the data on location basis. However, combined location analysis for attributes of ten genotypes that cut across the two locations was also done. The data generated were analyzed on location basis using SAS software (version 9.2) (SAS., 2008). Analysis of variance was performed on the data collected, on location basis and also as combined locations using the SAS GLM procedure. Both genotypes and locations were considered as fixed factor. The means of the genotypes were separated using Fisher’s LSD0.05. Phenotypic correlation coefficients, genotypic, environmental and phenotypic variances and broad-sense heritability were calculated on location basis, as well as combined locations for each trait. Phenotypic correlation among traits was analyzed using the SAS Proc Corr procedure. The SAS Princomp procedure was used to carry out the principal component biplot analysis. Phenotypic variance, genotypic variance and broadsense heritability estimates were calculated using the expected means square method. The variance components (Ve, Vg and Vp) were computed from a linear function of the means squares of the ANOVA according to Burton and Devane (1953) as follow:
For single location analysis:
Ve = MSe, Vg = (MSg-MSe)/r, Vp = Vg+Ve, HB = Vg/Vp
Where:| Ve | = | Environmental variance component |
| MSe | = | Error mean square |
| Vg | = | Genotypic variance component |
| Vp | = | Phenotypic variance component |
| MSg | = | Genotypes mean square |
| r | = | Number of replicates |
| HB | = | Broadsense heritability |
For combined location analysis:
Vg = (MSg-MSe)/rl
where, l = locations Ve, Vp and HB were calculated using same formulae as for single location broad-sense heritability analysis.
RESULTS
The analysis of variance (ANOVA) of the four processing traits (starch content, dry matter, peel-loss and flour content) evaluated in two contrasting locations (Makurdi and Umudike) showed that there was significant genotypic variability for the traits in each location and across locations. At each location, very highly significant (p<0.0001) differences existed among the genotypes for all the traits (Table 1). The combined locations ANOVA (Table 2) also showed that all the traits were under the control of genetics, the location of cultivation, as well as Genotype-by-Location Interaction (GEI). The GEI variance was lower than genotypic variance of all the traits.
The means of the genotypes for each trait on location basis are presented in Table 3 (Makurdi) and Table 4 (Umudike). At Makurdi, starch content ranged between 14.02% (UMUSP/2) and 20.53% (NRSP/05/1B) with only NRSP/05/1B having >20% starch.
| Table 1: | Analysis of variance of four processing traits of twelve sweetpotato genotypes planted in Makurdi, Benue state and Umudike, Abia state in 2013 |
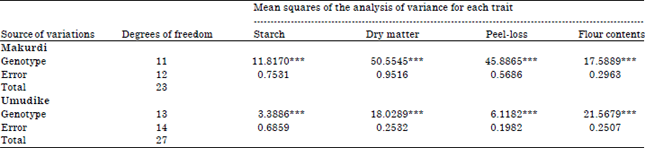 | |
| ***: p<0.0001 | |
| Table 2: | Mean squares of the combined analysis of variance of four sweetpotato processing traits across ten advanced sweetpotato breeding lines evaluated in two locations in 2013 |
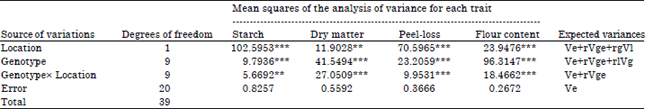 | |
| Ve: Environmental variance, Vg: Genotypic variance, Vge = Genotype×Environment variance, **: p<0.01, ***: p<0.0001 | |
| Table 3: | Means of sweetpotato processing traits of twelve sweetpotato genotypes planted in Makurdi, Benue state in 2013 |
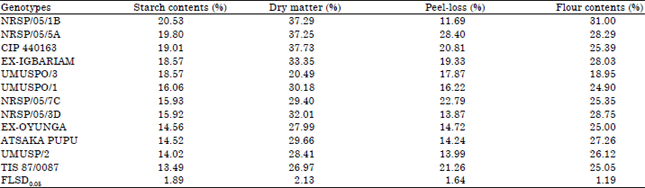 | |
| Table 4: | Means of sweetpotato processing traits of fourteen sweetpotato genotypes evaluated in Umudike, Abia state in 2013 |
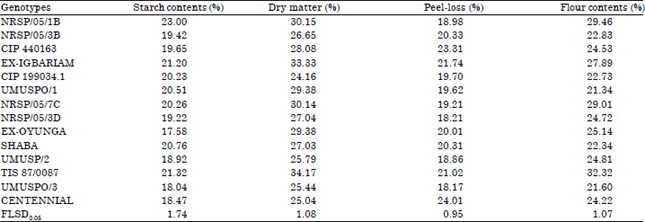 | |
For dry matter content, six genotypes-NRSP/05/1B, NRSP/05/5A, CIP 440163, UMUSPO/1, Ex-Igbariam and NRSP/05/3D, all had dry matter contents above 30%. The CIP 440163 had the highest dry matter of 37.73% while, CIP 440293 with 20.49% had the lowest dry matter content. Peel-loss had a range of 11.69-28.40%. Genotype NRSP/05/1B had the lowest peel-loss of 11.69%, while NRSP/05/5A, CIP 440163, NRSP/05/7C and TIS 87/0087 all had peel-loss > 20.0%. As for flour contents, NRSP/05/1B had flour content of 31.0% and was the only genotype with flour content up to 30%. NRSP/05/5A (28.0%), CIP 440163 (25.39%), Ex-Igbariam (28.03%), NRSP/05/7C (25.35%), NRSP/05/3D (28.75%), Ex-Oyunga (25.0%), Atsaka Pupu (27.26%), UMUSP/2 (26.12%) and TIS 87/0087 (25.05%0 all had at least 25% flour content.
At Umudike (Table 4), the starch content of the genotypes ranged between 18.47 and 23.00%, dry matter between 25.04 and 34.17%, peel-loss between 18.17 and 24.01% and flour content between 21.34 and 29.46%. NRSP/05/1B had the highest starch content of 23.00%. This was significantly (p<0.05) higher than the starch contents of the rest genotypes. However, six other genotypes had starch contents > 20%. For dry matter in the same location (Umudike), TIS 87/0087 with 34.17% had the highest dry matter. Other genotypes with dry matter content up to 30.0% are Ex-Igbariam (33.33%), NRSP/05/1B (30.15%) and NRSP/05/7C (30.14%). Peel-loss is a trait whereby the smaller the value, the better the genotype. Four genotypes had peel-loss of about 18%, which is the least. Centennial had the highest peel-loss of 24.01%, meaning almost one-fourth of the root weight was lost to peeling. Flour content is a processing attribute that is important to sweetpotato processors. TIS 87/0087 had the highest and was the only genotype with flour content that was up to 30% (32.32%).
Combined analysis was performed to study the mean performance of each genotype across locations for each of the processing traits; to show location effect on each trait across genotypes; as well as to identify which genotype at which location had the best trait expression. The results of the combined analyses are presented in Table 5-8. Table 5 shows the means of genotype, location and genotype-by-location interaction for starch content. Among the genotypes, NRSP/05/1B had the highest mean starch of 21.77% across locations. Other genotypes had significantly (p<0.05) lower mean starch. For location effect, Umudike supported more (p<0.05) mean starch accumulation (19.87%) compared to Makurdi (16.66%). Over all, NRSP/05/1B at Umudike gave the highest starch content of 23.0%, followed by TIS 87/0087 at Makurdi with 21.32%. TIS 87/0087 at Makurdi had the lowest starch of 13.49%. For mean dry matter accumulation by each genotype across locations (Table 6), NRSP/05/1B (33.72%), Ex-Igbariam (33.34%), CIP 440163 (32.91%) and TIS 87/0087 (30.57%) all had mean dry matter > 30%, while other genotypes had lower dry matter contents.
| Table 5: | Combined mean of starch contents (%) across ten advanced sweetpotato breeding lines and two locations measured in 2013 |
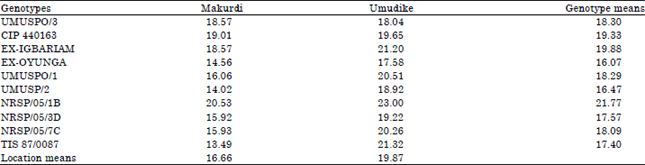 | |
| FLSD0.05 for comparing location means = 0.60, FLSD0.05 for comparing genotype means = 1.34, FLSD0.05 for comparing GX E means = 1.90 | |
| Table 6: | Combined mean of dry matter contents (%) across ten advanced sweetpotato breeding lines and two locations measured in 2013 |
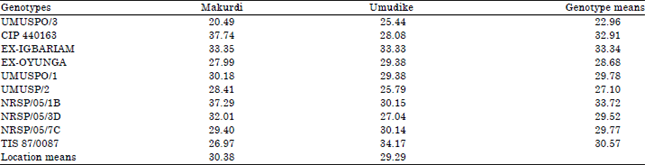 | |
| FLSD0.05 for comparing location means = 0.49, FLSD0.05 for comparing genotype means = 1.10, FLSD0.05 for comparing GX E means = 1.56 | |
| Table 7: | Combined mean of peel-loss (%) across ten advanced sweetpotato breeding lines and two locations measured in 2013 |
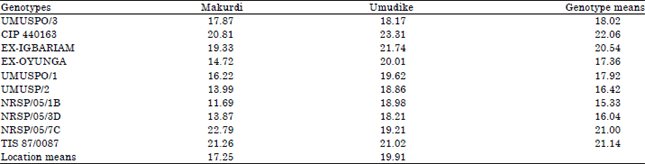 | |
| FLSD0.05 for comparing location means = 0.40, FLSD0.05 for comparing genotype means = 0.89, FLSD0.05 for comparing GX E means = 1.26 | |
The variation in dry matter makes selection among the genotype for processing possible. The savannah ecology of Makurdi resulted in higher mean dry matter accumulation compared to mean dry matter contents across genotypes at Umudike, rainforest ecology. CIP 440163 at Makurdi (37.74%) and NRSP/05/1B at Makurdi (37.29%) had the highest dry matter.
Table 7 presents the combined mean of peel-loss (%) across the two locations for each genotype, as well as peel-loss for each genotype at a specific location. Genotype CIP 440163 had the highest mean peel-loss of 22.06% across locations which was significantly higher (p<0.05) than others. This is a significant waste that will lead to reduced roots for further processing this genotype. Lowest mean peel-loss of 15.33% was recorded by NRSP/05/1B. For reasons unknown, genotypes at Umudike had higher (p<0.05) mean peel-loss of 19.91% than the 17.25% recorded in Makurdi. In general, NRSP/05/1B at Makurdi had the lowest tissue loss due to peeling of 11.69%, while CIP 440163 at Umudike with peel-loss of 23.31% had the highest loss to peeling.
| Table 8: | Combined mean of flour contents (%) across ten advanced sweetpotato breeding lines and two locations measured in 2013 |
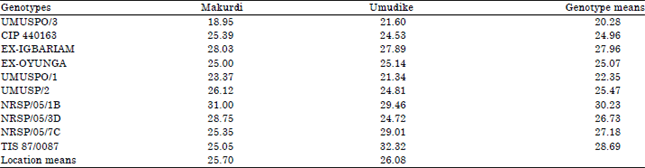 | |
| FLSD0.05 for comparing location means = 0.39, FLSD0.05 for comparing genotype means = 0.76, FLSD0.05 for comparing GX E means = 1.08 | |
| Table 9: | Correlation studies among four processing traits of sweetpotato planted in Makurdi, Benue state and Umudike, Abia state in 2013 |
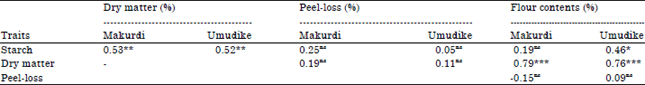 | |
| Twelve advanced sweetpotato breeding lines (genotypes) were involved in the trial at Makurdi, while 14 genotypes were involved in the trial at the Umudike location, ns: Not significant (p<0.05), *: p<0.05, **: p<0.01, ***: p<0.0001 | |
| Table 10: | Combined phenotypic correlation coefficients (r) among four processing traits of sweetpotato across ten genotypes and two locations in 2013 |
 | |
| Ten genotypes that were common to both locations were used for the analysis. Analysis for each trait was replicated three times, ns: Not significant, *: p<0.05 | |
For flour content (Table 8) and as expected due to high dry matter, NRSP/05/1B had the highest mean flour content of 30.23%. UMUSPO/3, an orange-fleshed sweetpotato, had the lowest mean flour content of 20.28% across the two locations. For flour content in each location across genotypes, there was no significant difference (p>0.05) between Umudike and Makurdi. However, only genotypes TIS 87/0087 at Umudike (32.32%) and NRSP/05/1B at Makurdi (31.00%) had flour contents >30%. Flour content of 30% and above will enhance the adoption of sweetpotato genotypes for commercial processing.
Correlation studies among the four processing traits were carried out at each location and as combined locations to determine the relationship that exist among the traits as expressed by the genotypes. At both Umudike and Makurdi, dry matter content had significant and positive correlation with starch content (p<0.01) and flour content (p<0.0001) (Table 9). Starch showed a positive and significant (p<0.05) relationship with flour content only at Umudike. Peel-loss had no relationship whatsoever with other traits. When the data were combined across locations, same trend of positive and significant (p<0.05) relationships were found between dry matter and starch contents and between dry matter and flour contents (Table 10). However, starch content showed significant relationship with peel-loss contrary to what was observed in single location correlation analyses. It was observed that correlation coefficients (r) and levels of significance (p) were reduced in the combined location analysis compared to those of single locations.
Principal component biplot was carried out to visually show the relationship between genotype(s) and the processing traits in a manner that enhances better selection.
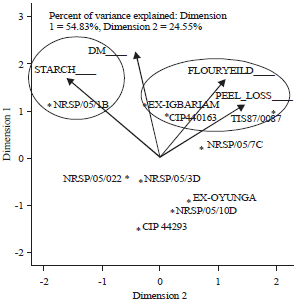 | |
| Fig. 1: | Principal component analysis biplot of ten sweetpotato genotypes and four processing traits measured in two locations in 2013. The biplot explains 79.38% of the total variance among genotypes for the four processing traits |
| Table 11: | Heritability estimates of important processing traits of sweetpotato evaluated in Makurdi, Umudike and combined locations in 2013 |
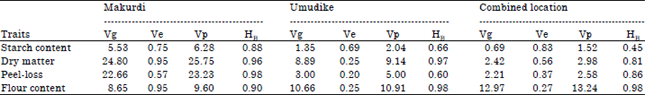 | |
| Vg: Genotypic variance, Ve: Environmental variance, Vp: Phenotypic variance HB: Broadsense heritability | |
Figure 1 shows the principal component biplot of the combined locations for genotypes and traits. The biplot shows that genotypes NRSP/05/1B, Ex-Igbariam, CIP 440163 and TIS 87/0087 all have elevated expressions of high contents of starch, dry matter, peel-loss and sweetpotato flour. High starch content and high dry matter were highly related to genotype NRSP/05/1B. While Ex-Igbariam, CIP 440163 and TIS 87/0087 also expressed high dry matter, the three genotypes exerted more influence on flour content and peel-loss. The other genotypes-NRSP/05/022, NRSP/05/10D, NRSP/05/3D, CIP 440293 and Ex-Oyunga had average to low values of the traits. Incidentally, all the three orange-fleshed genotypes (NRSP/05/022, CIP 440293 and Ex-Oyunga) involved in this study belong to this group.
The genotypic (Vg), environmental (Ve) and phenotypic (Vp) variances, as well as the broad-sense heritability estimates of the four processing traits at each location and combined locations were presented in Table 11. For all the traits in each location and across locations, Vg was higher than Ve. At Umudike and Makurdi, the broad-sense heritability (HB) estimates were either moderate or high and ranged between 0.60 and 0.98. Heritability estimate for starch content at Makurdi was 0.88 while, it was 0.66 at Umudike. For dry matter, HB estimates were 0.96 and 0.97 at Makurdi and Umudike, respectively and 0.98 and 0.60 for peel-loss at Makurdi and Umudike respectively. While, HB for flour content at Makurdi was 0.90, it was 0.98 for the trait at Umudike. For combined location analysis, HB for starch content was moderate at 0.45, while HB for dry matter was 0.81. Peel-loss (0.86) and flour content (0.98) also had high heritability estimates.
DISCUSSION
The availability of variability in desirable traits among genotypes in a breeder’s germplasm is critical for breeding progress as effective selection depends on the presence significant genetic variability. According to Collins (1990), there is tremendous genetic variability among sweetpotato genotypes for many traits. This confirms the wide variability observed for the processing traits (dry matter, starch and flour contents and peel-loss) in this work.
Determining the expression of desirable traits in more than one location helps to study the presence of Genotype-by-Environment Interaction (GEI), the differential response of genotypes at different locations for desirable traits (Acquaah, 2007). Significant GEI were observed for all the four traits in this work. Belehu (2003) and Gruneberg et al. (2005) had observed significant GEI for root dry matter and starch contents among sweetpotato genotypes. Significant GEI depicts that genetic materials must be evaluated for desirable traits for many years in the environments where such materials will be released and this quite often delays the varietal selection process (Claiskan et al., 2007).
High dry matter (Tumwegamire et al., 2004) and starch contents (Dansby and Bovell-Benjamin, 2003; Chen et al., 2003b) have been reported as very important attributes that influence sweetpotato consumers’ preferences and industrial utilization. Mwanga et al. (2003) put the least acceptable dry matter content for sweetpotato processing at greater than 30%. Six of the advanced lines had root dry matter greater than 30%. Various authors had reported different dry matter, starch content and flour content ranges for sweetpotato. The range of dry matter observed in this work in the two locations was within the dry matter range of 15.0-35.0% reported by Cervantes-Flores et al. (2011) and 20.0-37.8% reported by Karuri et al. (2009). The starch content of the genotypes used in this work was close to the 17.58-22.00% reported by Afuape et al. (2014). So, also was the flour content within the range reported by Afuape et al. (2014). The wide range between the genotypes for these processing traits gives room for the selection and official release of genotypes that combine these processing traits with good agronomic attributes. Selection of parent lines among the genotypes used is also possible for recombination.
The desire of a plant breeder is to know the extent of relationship between important traits. The significant and positive correlation that exists between dry matter and starch content and between dry matter and flour contents, means that both starch and flour contents can be simultaneously improved along with dry matter. These relationships had also been reported by Cervantes-Flores et al. (2011) and Afuape et al. (2014).
Heritability is a measure of the correspondence between phenotypic values and breeding values. High heritability estimate suggests that superior parents tend to give the best progeny (Bernardo, 2002) and so is used by breeders as prediction tool to predict the performance of the offspring from that of their parents.
High heritability estimates were observed for dry matter in both locations as well as combined locations (0.81-0.97). Moderate to high heritability estimates had been reported by such authors as Jones (1986) (HN = 0.65) and Courtney (2007) (HN = 0.92). The moderate heritability recorded for starch contents had previously been observed also by Afuape et al. (2014). The moderate value for starch suggests that the trait is under additive gene action. This is corroborated by the findings of Dai et al. (1988) that starch content in sweetpotato was under the control of additive gene effect. The moderate to high heritability estimates of all the four traits point to the prospects of a rapid genetic gain in breeding for improved dry matter, starch and flour contents using these genetic materials.
Having six genotypes with dry matter >30% and heritability estimates of 0.81 (combined locations), 0.96 (Makurdi location) and 0.97 (Umudike location) give room for the possibility of developing new sweetpotato varieties with elevated dry matter from the genetic materials evaluated as parent lines. Developing varieties that combine high dry matter, high starch and high flour contents will not only enhance the processing of sweetpotato, it will also enhance the consumption of the products from such varieties through the development of consumer-preferred products. This is because starch is a strong determinant of the physico-chemical, rheological and textural characteristics of food (Ezeocha et al., 2013). As a functional food biopolymer, it influences the quality of the final food products (Brunnschweiler et al., 2005). The moderate heritability of starch points to the fact that many genes are probably involved in starch biosynthesis and the environment has more influence on the expression of important genes in the sweetpotato starch biosynthetic pathway. In cassava, ADP-glucose pyrophosphorylase, starch synthase and starch-branching enzymes were identified as important in the cassava starch biosynthetic pathway (Saelim, 2008). A recurrent selection method, which is often used for improving traits with low heritability, will be adequate to improve starch content in sweetpotato.
CONCLUSION
This study has been able to show the extent of variability that exist among the genotypes, the expression of the important processing traits as influenced by location of cultivation, how heritable or otherwise the traits are, as well as the relationship that exist among these traits. Attempts had been made to link all of these, showing the implication of this study to breeding for sweetpotato utilization. The study also supplies the information that point towards the effective use of sweetpotato germplasm and selection methods for developing varieties that are adequate for processing.
ACKNOWLEDGMENTS
This study emanated from the Alliance for Green Revolution in Africa (AGRA) -sponsored Sweetpotato Breeding Project (2010 PASS 002) at the National Root Crops Research Institute (NRCRI), Umudike, Nigeria. The field superintendents, Mr. Nwankwo, C.K. and Mrs. Ogbo, E. and the laboratory staff who helped during the laboratory analyses are acknowledged for their efforts. The Sweetpotato Programme of the NRCRI, Umudike is also acknowledged for the support in carrying out this work.
REFERENCES
- Afuape, S.O., I.I.M. Nwankwo, R.M. Omodamiro, T.N.C. Echendu and A. Toure, 2014. Studies on some important consumer and processing traits for breeding sweet potato for varied end-uses. Am. J. Exp. Agric., 4: 114-124.
Direct Link - Brunnschweiler, J., D. Luethi, S. Handschin, Z. Farah, F. Escher and B. Conde‐Petit, 2005. Isolation, physicochemical characterization and application of yam (Dioscorea spp.) starch as thickening and gelling agent. Starch‐Starke, 57: 107-117.
CrossRefDirect Link - Burton, W.G. and E.H. Devane, 1953. Estimating heritability in tall fescue (Festuca arundinacea) from replicated clonal material. Agron. J., 45: 478-481.
CrossRefDirect Link - Claiskan, M.E., E. Erturk, T. Sogut, E. Boydak and H. Arioglu, 2007. Genotype x environment interaction and stability analysis of sweetpotato (Ipomoea batatas) genotypes. New Zealand J. Crop Hortic. Sci., 35: 87-99.
CrossRefDirect Link - Cervantes-Flores, J.C., B. Sosinski, K.V. Pecota, R.O.M. Mwanga and G.L. Catignani et al., 2011. Identification of quantitative trait loci for dry-matter, starch and β-carotene content in sweetpotato. Mol. Breed., 28: 201-216.
CrossRefDirect Link - Chen, Z., H.A. Schols and A.G.J. Voragen, 2003. Physicochemical properties of starches obtained from three varieties of Chinese sweet potatoes. J. Food Sci., 68: 431-437.
CrossRefDirect Link - Chen, Z., H.A. Schols and A.G.J. Voragen, 2003. Starch granule size strongly determines starch noodle processing and noodle quality. J. Food Sci., 68: 1584-1589.
CrossRefDirect Link - Dai, Q., R. Qiu, P. Xu and Y. Xie, 1988. Genetic parameters of quantitative traits and breeding strategy for high starch content and high yield in sweet potato. Scientia Agricultura Sinica, 21: 33-38.
Direct Link - Ezeocha, V.C., E. Oti, S.C. Chukwuma, C. Aniedu and O.N. Eke-Okoro, 2013. Evaluation of the physico-chemical properties of twelve D. rotundata land races grown in south-east agro-ecology. Adv. Applied Sci. Res., 5: 21-25.
Direct Link - Fawole, O.P., 2007. Constraints to production, processing and marketing of sweet potato in selected communities in Offa Local Government Area, Kwara State, Nigeria. J. Hum. Ecol., 22: 23-25.
Direct Link - Gruneberg, W.J., K. Maniique, D. Zhang and M. Hermann, 2005. Genotype x environment interactions for a diverse of sweet potato clones evaluated across varying eco-geographic conditions in Peru. Crop. Sci., 45: 2160-2171.
Direct Link - Karuri, H.W., E.M. Ateka, R. Amata, A.B. Nyende and A.W.T. Muigai, 2009. Characterization of Kenyan sweet potato genotypes for SPVD resistance and high dry matter content using simple sequence repeat markers. Afr. J. Biotechnol., 8: 2169-2175.
Direct Link - Dansby, M.Y. and A.C. Bovell‐Benjamin, 2003. Production and proximate composition of a hydroponic sweet potato flour during extended storage. J. Food Process. Preservation, 27: 153-164.
CrossRefDirect Link - Mwanga, R.O.M., B. Odongo, G. Turyamureeba, A. Alajo and G.C. Yencho et al., 2003. Release of six sweetpotato cultivars ('NASPOT 1'to'NASPOT 6') in Uganda. HortScience, 38: 475-476.
Direct Link - Tumwegamire, S., R. Kapinga, D. Zhang, C. Crissman and S. Agili, 2004. Opportunities for promoting orange-fleshed sweetpotato as a mechanism for combat vitamin-A deficiency in Sub-Saharan Africa. Afr. Crop Sci. J., 12: 241-252.
Direct Link