
G. Fathi
Department of Agronomy, Ramin Agricultural and Natural Resources University, Mollasani, Ahwaz, Iran
Journal of Agronomy
Year: 2006 | Volume: 5 | Issue: 2 | Page No.: 267-277







ABSTRACT
Grain yield and protein in wheat are determined by the plant efficiences in partitioning dry matter and Nitrogen (N) to the grain. The interaction between post-anthesis water stress and nitrogen rate was examined in six wheat cultivars. Plant were grown in a glasshouse at 2 rates of N under well-watered conditions until 7 days affer ear emergence, when the stress treatment was started. Yield and Grain Protein Concentration (GPC) responses and changes in the dry matter and N content of the straw and grain in the main stem and tillers were examined. Nitrogen increased grain yield in all cultivars except Fong, with Atila and Falat being the most responsive. Yields at the low N rate did not reduce by post-anthesis stress, but large reductions occurred at the high N rate in all cultivars; the yield of Falat, Showa and Atila were most affected. At the low N rate, stress did not significantly affect kernel weight and GPC, but kernel weight declined and GPC increased at the high N rate. Tillers produced smaller grain with a lower GPC than main stem. The responses to N and water stress and the different sensitivities of cultivars to stress, were largely due to the effects of the treatments on the growth of the tillers. Net remobilisation of dry matter was increased by stress but not by N treatment and the amount remobilised varied between genotypes. Post-anthesis stress increased the N content per kernel and net remobilisation of N at the high N rate. Although genotypes differed in the net amount of N remobilised and in the N harvest index, there was little variation in GPC between cultivars. Results showed that reductions in yield and kernel weight and increases in GPC from post-anthesis stress can be greater when plants are grown at a high rate of N than when the supply of N is limited. The different responses to stress and N among the six wheat cultivars were associated, in part, with the pattern of tiller development. However, there appeared to be differences in the sensitivity of grain filling to stress independent of the responses in tillering. While the net remobilisation of dry matter and N differed between cultivars, the amounts did not appear to be related to differences in GPC.
PDF Abstract XML References
How to cite this article
G. Fathi, 2006. Post-anthesis Water Stress and Nitrogen Rate Effects on Dry Matter and Nitrogen Remobilisation in Wheat Cultivars. Journal of Agronomy, 5: 267-277.
DOI: 10.3923/ja.2006.267.277
URL: https://scialert.net/abstract/?doi=ja.2006.267.277
DOI: 10.3923/ja.2006.267.277
URL: https://scialert.net/abstract/?doi=ja.2006.267.277
INTRODUCTION
The area of farming in Iran is more than 12 million ha, which are mostly devoted to wheat production (6.2 million ha). Grain Yield (GY) and Grain Protein Concentration (GPC) in wheat are determined by the plant efficiencies in partitioning dry matter and Nitrogen (N) to the grain. The profitability of wheat production, however is affected largely by application N fertilizer increasingly, farmers are applying N fertilizer wheat and the yield responses achieved with applications of N fertilizer are generally higher and more consistent than those obtained in barley (Brich and Long, 1990; Ehdaie and Waines, 2001). Nevertheless, applying N to wheat is not without risk. The Mediterranean-type environment of south of Iran is characterised by declining rainfall in spring and increasing evaporation rates, which often result in water deficits sufficiently great to reduce plant growth. Nitrogen applied at sowing can promote tillering and dry matter production (Brich and Long, 1990; Palta and Fillery, 1995a), but if spring rainfall is below average the yield response to may be limited by exhaustion of soil moisture reserves and GPC may increase. High concentrations of N in plant tissues for the addition of N fertilizer may also alter a plant’s sensitivity to increasing moisture deficits (Morgan, 1986; van Keulen and Stol, 1991), resulting in plants being less tolerant of water deficits than plants with low tissue N concentrations. These studies suggest that the use of N fertilizer may increase the risk of reductions in growth from moisture stress, which may adversely affect grain quality.
High grain yield and GPC are important for grain quality of wheat, and both of these characteristics depend on Dry Matter (DM) and N remobilized from the leaves and stems to the grain. However, their respective contributions to yield and grain N content vary considerably (Bidinger et al., 1977; Pheloung and Siddique, 1991; Palta et al., 1994). Remobilization of DM may help to maintain grain growth when post-anthesis photosynthesis is reduced (Solfied et al., 1977; Papakosta and Gagianas, 1991). Studies using post-anthesis desiccation to simulate drought have demonstrated genotypic variation in the reduction in kernel weight under stress in wheat (Blum et al., 1983; Turner and Nicolas, 1987; Hossain et al., 1990), which suggests genotypic difference in the ability to remobilized dry matter to maintain grain growth. If high N rates exacerbate the adverse effects of post-anthesis drought, the ability of a plant to remobilizedd DM to maintain grain growth may be an important characteristic for wheat cultivars. Under the conditions normally experienced in south of Iran, the N requirement of the developing grain is satisfied largely by the mobilization of N from vegetative tissues (Dalling et al., 1976), although in situations where terminal drought is not serves, N uptake during grain filling may account for a significant proportion of the grain N at maturity (Spiertz and Ellen, 1978; Gregory et al., 1981). It is desirable that wheat cultivars are able to maintain GY and GPC within a specified range across environments and this may depend on developing cultivars that maintain grain growth under stress and/or remobilise high amounts of N to the grain.
As well as affecting the amount of N and DM remobilised, N and post-anthesis water stress also alter their partitioning between the main stem and tillers. With increasing levels of stress, DM and N are translocated from the later formed tillers to the main stem and first-formed tillers (Chafai et al., 1992; Kobata et al., 1992), but the amount remobilizedd and translocated to the main stem is influenced by the stage of development of the tillers at the time when sets occurs (Chafai et al., 1992). In studies on responses to plant density in wheat, Turner et al. (1994) found that the kernel weight of grain from tillers was less than that on the main stem and this was associated with the number of tillers produced by the plant and rate of development of the tillers relative to the main stem. Therefore, the relative stages of development of tillers on a plant (tiller synchrony) may affect the amount of remobilization of DM and N from the tillers and the partitioning between main stem and tillers. The current study examined the effect of post-anthesis water stress on the yield and GPC of 6 wheat cultivars grown at 2 N rates. The cultivars were chosen because they had been used previously in field experiments and they represented a range of wheat cultivars that has been grown in south of Iran. It also examined whether those cultivars more responsive to N under favourable conditions were more sensitive to post-anthesis drought. The importance of the interaction between the main stem and the tillers in these responses was evaluated by measuring the DM and GY response of the main stem and the tillers separately.
MATERIALS AND METHODS
The experiment was conducted in an air-conditioned glasshouse in November 2002 under normal day lengths. Six cultivars of wheat (Fong, Atila, Falat, Atrak, Star and Showa) was grown at 2 rates of N and kept either well watered or stressed after anthesis as described below. To allow an initial harvest so that change in growth and N distribution could be measured, extra plants were grown the 2 N rates and kept well watered up to ear emergence, when they were harvested. These posts were arranged randomly with the other post of the experiment. The plants were grown in 16 cm-diameter pot lined with plastic bags containing in 2.4 kg of steam sterilised soil mix (1:1 sand: loam mix) to which no N was added but which was adequate for the other plant nutrients. The potting mix had a pH of about 7.5 and the concentration of mineral N (ammonium-N plus nitrate-N) in the soil at sowing was 9.4 μg m-1. Four seed per pot were sown on 19 November 2002 and seedling were thinned to 2 plants per pot after emergence.
Nitrogen was applied as a solution of ammonium nitrate (34% N) at 274 and 540 mg of ammonium nitrate per pot, respectively, to the low and high N treatments in 3 equal applications: at the 2-leaf stage (growth stage 12, Zadokes et al., 1974) at tillering (GS21) and at stem elongation (GS31).
Plants were watered frequently until emergence of the ear on the main stem (GS 58). At the start of the experiment, 4 pots containing soil, but with no plastic bag liners, were saturated with water, covered with black plastic and allowed to drain to a constant weight. The gravimetric moisture content of the soil in the pots. Pots was subsequently rewatered to this weight. Two watering treatments (well watered and water-stressed) were imposed 7 days after ear emergence. Pots in the well-watered treatment were maintained at field capacity by frequent weighing and the addition of water to return them to field capacity, whereas pots in the water-stressed treatment received half the quantity of water supplied to the well-watered pots at each watering.
Plants grown at the 2 N rates were harvested 10 days after the main stem ear had emerged (H1) and at maturity (H2). Although H1 occurred after the start of the stress treatment, no visible signs of stress were evident at this time. At each harvest the numbers of tillers and ears per pot were counted, the roots were washed from the soil and the shoots were divided into main stem and tillers. Shoot, root and grain from each replicate were oven dried at 80°C for 48 h and weighted and the N concentration was determined by kjeldahl analysis. Grain N concentration was multiplied by 6.25 to grive the GPC. The number of kernels from the main stem and the tillers was counted at maturity and main stem and the tillers was counted at maturity and the average kernel weight derived from the grain weight and the kernel numbers. The N content per kernel was estimated from the grain N yield (grain N concentrationxgrain yield) and the number of kernels per m2. Net Dry Matter Remobilized (DMR) and net N remobilized (NR) from the straw between ear emergence (H1) and maturity (H2) were calculated using the following equations:
Where non-grain DM=(total DM-grain dry weight) and non-grain total N = (total N-grain N). Both DMR and NR are used as estimates of the amounts of dry matter and N remobilized in the experiment, but we recognize that DMR does not take account of respiratory losses or the contribution from post-anthesis photosynthesis, or that NR accounts for losses of N from plant tissue by volatilization.
The distribution of DM and N between the main stem and the tillers was quantified by calculating the proportion of total plant dry matter, grain yield and plant N content present in the main stem. For example a dry matter ratio of 0.30 means that the dry matter of the main stem represented 30% of the total plant dry weight. The experiment was designed as a factorial randomized complete block with 4 replicates, at the first harvest the treatments were 6 cultivarsx2 N ratesx2 stress treatments. For the analysis of variance, the treatments were considered to be fixed (Steele and Torrie, 1960) and the data were analyzed using SAS methods.
RESULTS
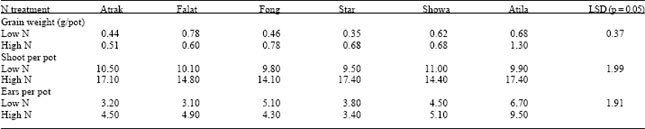
Growth at 10 days after anthesis (H1): The average concentration of N in the shoots at the lower rate of N (1.1%) indicated that the plants were N-deficient. Consequently, the additional N substantially increased the growth, grain weight and GPC of all cultivars. Nitrogen significantly increase average shoot dry matter from 9.4 to 13.3 g/pot and average root dry matter from 3.4 to 4.2 g/pot. There were no significant differences between cultivars in root or shoot dry weight, nor were there significant cultivarxN interactions. Grain weight per pot increased significantly with additional N only in Atila (Table 1) and this was due largely to the increased yield from the tillers.
Total shoot and ear numbers were increased at the high N rate. At the low rate of N there were no significant difference in the number of shoots and ears per pot among cultivars, but at the high rate of N, Atrak and Star produced significantly more tillers than the other cultivars. Ear emergence had not occurred on many of the tillers at this stage and so ear number were substantially lower than total shoot number, although the general trends in shoot and ear numbers were similar. Comparisons of the number of shoots with the number of ears indicated that there were differences in the synchrony of ear emergence. Atila, as well as producing a large number of tillers had a larger proportion of these producing ears, suggesting a more synchronous pattern of ear emergence than Atrak or Star, both of which produced numbers of tillers similar to Atlia but fewer ears. The extra N increased tillering and consequently, the contribution of the main stem to total plant dry matter declined from 38 to 27%. At this stage 53% of the total plant grain weight was from the main stem and there was no significant effects of N on this distribution.
There were no significant differences between cultivars in GPC, but the higher rate of N increased no significant interaction between N and cultivar for GPC. Grain form the tiller ears had 1% higher GPC, on average, than grain from the main stem.
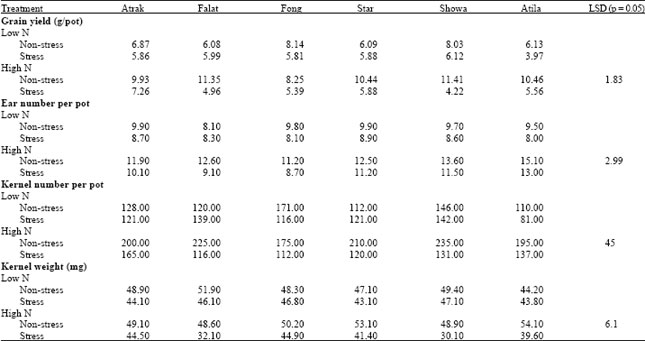
Maturity: In general, post-anthesis stress reduced grain yield and kernel weight and increased GPC, but these effects of stress were greater when plants were grown at the higher N rate (Table 2). For example, stress significantly reduced grain yield in all cultivars at the high rate of N, with Falat, Showa and Atila being particularly sensitive, but at the low N rate only the yield of Fong, Showa and Atila were significantly reduced by stress. The greater sensitivity of Falat, Showa and Atila to drought at the higher level of N mirrored the changes in both kernel number and kernel weight (Table 2). However, the reduction in kernel weight of the these cultivars appeared to be particularly sensitive, because their kernel weight when grown under stress at high N was significantly less than when grown under stress at low N. When all data were combined, grain yield and the total post-anthesis dry matter accumulation were positively correlated (r = 0.85, p<0.01, n = 24). The main factors affecting GPC were N treatment and post-anthesis stress. Cultivar was relatively less important and differences were mainly observed in the protein concentration of the grain from the tillers (Table 3). Post-anthesis stress did not increase GPC at the low N level but increased it substantially at the high N level (Table 3).
| Table 1: | Effects of N application rate on grain weight, shoots number and ears number in 6 wheat cultivars, 10 days after anthesis on the main stem (the data shown are for well-watered plants only) |
 | |
| Table 2: | Effects of post-anthesis water stress on the grain yield, ear number, kernel number and kernel weight of 6 wheat cultivars grown at two rates of N |
 | |
| Table 3: | Effects of post-anthesis water stress, averaged across cultivars, on the grain protein concentration, N content per kernel and N harvest index in 6 wheat cultivars grown at 2 N rates |
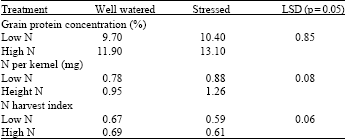 | |
The N content per kernel showed responses to post-anthesis stress and N treatment similar to GPC (Table 3), suggesting that the responses measured in GPC were associated with increased movement of N into the grain and not only with the reduction in kernel weight (Table 3). The average N content per kernel was 0.66, 0.62, 0.74, 0.70, 0.60 and 0.70 mg (LSD p = 0.05, 0.073) for Atrak, Falat, Fong, Star, Showa and Atila, respectively. High N reduced the N harvest index (NHI) and water stress reduced NHI at the high N rate but not at the low N rate (Table 3).
Net remobilization of dry matter and N: Nitrogen treatment and water stress had quite different effects on the remobilization of dry matter and N per plant. When kept well watered, there was virtually no net remobilization of dry matter per plant from the straw because of the accumulation of dry matter in the tillers on the roots, but net remobilization increased significantly under straw, especially in the tillers and roots (Fig. 1). As a consequence of this and of the reduction in grain yield from stress, the amount remobilized as a proportion of yield increased (Fig. 1). The main stem accounted for at least 50% of the total straw dry mater remobilized and the loss of dry matter from the roots under stress was similar to that lost from the main stem. Nitrogen rate had no significant effect on the amount of dry matter remobilizeddd (Fig. 1). In contrast, stress did not significantly affect the amount of N remobilised from the main stem, tillers and roots, but there were significant increases in net remobilisation at the high rate of N (Fig. 1). Although the loss of N from the roots was increased at the higher N level, the amount as a proportion of the root N at H1 were similar (37 v. 39%).
| Table 4: | Effects of post-anthesis water stress on the proportion of total dry matter and grain yield derived from the main stem in 6 wheat cultivars growth at 2 N rates |
 | |
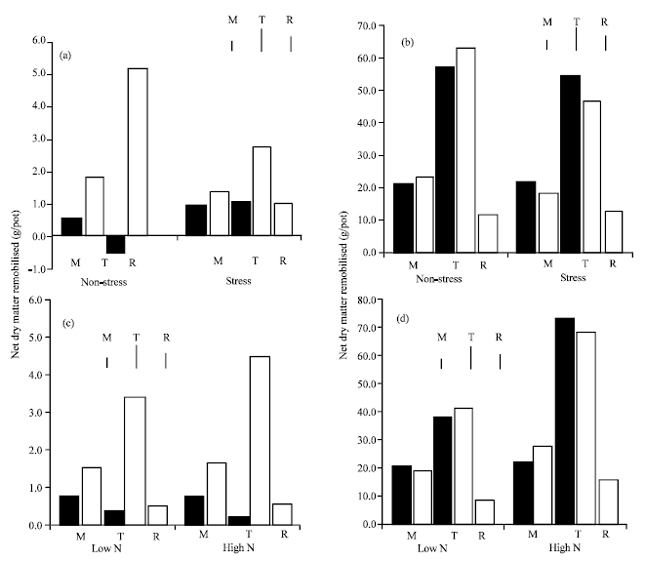 | |
| Fig. 1: | Effects of post-anthesis stress (a and b) and N treatment (c and d) on: (a, c) the net remobilisation of dry matter from the shoot (solid bars), the increase in grain yield (shaded bars) and the net remobilisation of dry matter from the root (open bars) and (b, d) net remobilisation of N (solid bars), the increase in grain N content (shaded bars) and the net remobilisation of N from the root (open bars) between H1 and maturity. Main stem (M), tillers (T) and roots (R) are indicated and each value is the average of 6 cultivars. Vertical lines are the LSD (p = 0.05) |
Also in contrast to net dry matter remobilization, tillers contributed much more to the total shoot remobilization of N than the main stem: whereas more than half of the loss of straw dry matter was measured in the main stem, only 25-35% of the loss of N was measured in the main stem.
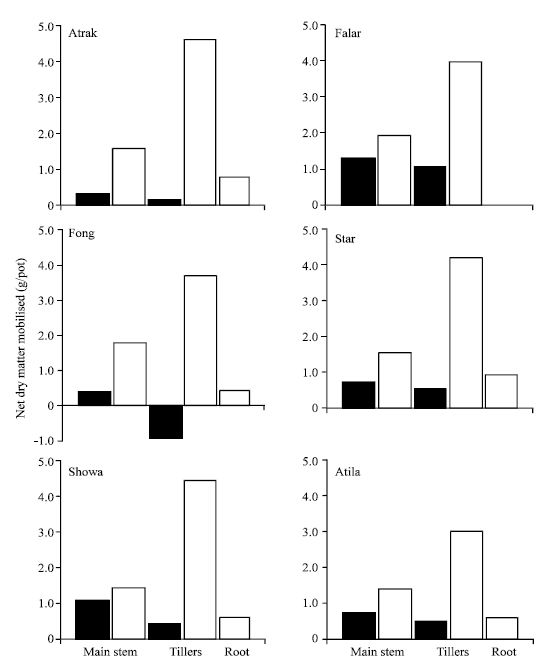 | |
| Fig. 2: | Remobilisation of dry matter (solid bars) and the increase in grain yield (shaded bars) in the main stem, tillers and roots (open bars) of six cultivars of wheat between H1 and maturity. The value of each cultivar is averaged over the N and water stress treatments. The LSD (p = 0.05) values for remobilisation of dry matter in the main stem, tillers and roots are 0.49, 1.44 and 0.78 g/pot, respectively |
Cultivar differences in the net dry matter remobilised from the whole shoot ranged from a net grain in straw weight of 0.55 g/pot in Fong, due to an accumulation of dry matter in the tillers, to a loss of 2.30 g/pot in Falat (Fig. 2). The net amount of dry matter remobilized relative to the increase in grain dry matter averaged 26% and did not differ significantly among cultivars. There were no significant differences among cultivars in the reduction in root dry matter post-anthesis, which averaged 0.51 g/pot, or about 14% loss of the root dry matter at H1.
The total net remobilization of N from the shoot in Falat, Star and Atila was about 80-90 mg/pot (Fig. 3), which was significantly greater than the net amount remobilised in the remaining cultivars (about 60-70 mg. pot). This difference was due largely to differences in the loss of N from the tillers.
As proportion of the increase in plant grain N content, the net amount of N remobilised averaged 107% but ranged from 79% (Fong) to 162% (Atlia).
Contributions of the main stem and tillers: The greater proportion of total plant yield was derived from the tiller ears at both N rates (Table 4, Fig. 4) and the responses of the whole plant to the N and stress treatments largely reflected the responses of the tillers. When grown under a high N rate and without sever moisture stress, the grain yields of Falat, Star, Showa and Atila were high (Fig. 4).
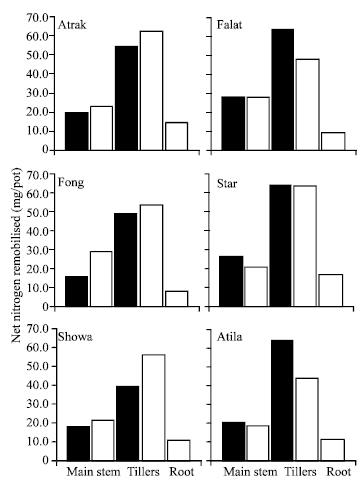 | |
| Fig. 3: | Remomilisation of N from the shoot (solid bars) and the increase in grain N in the main stem and tillers (shaded bars) and in the roots (open bars) of 6 cultivars of wheat between H1 and maturity. The value of each cultivar is averaged over the N and water stress treatments. The LSD (p = 0.05) values for remobilisation of N in the main stem, tillers and roots are 5.0, 12.7 and 1.0 mg/pot, respectively |
The large reductions in the yield of Falat and Showa at the high N rate were due to reductions in kernel numbers, particularly from the tillers (Fig. 4) and in the kernel weight from the tillers (Fig. 5). Thus, the proportion of grain yield derived from the main stem increased substantially (Table 4). A similar trend was evident in Atila, although the number of kernels derived from the tillers of Atila was not reduced as much as that of Falat and Showa and the reduction in kernel weight was relatively more important in determining the yield response of Atila. The contribution of the tillers to the total plant grain yield in Atila remained higher than in Falat and Showa. The yield of Atrak was not reduced as much by stress as that of the other cultivars at the high N rate (Table 2), largely because of its ability to maintain a relatively high yield on the tillers (Fig. 4), therefore, the distribution of yield between main stem and tillers was not significantly affected by stress (Table 4).
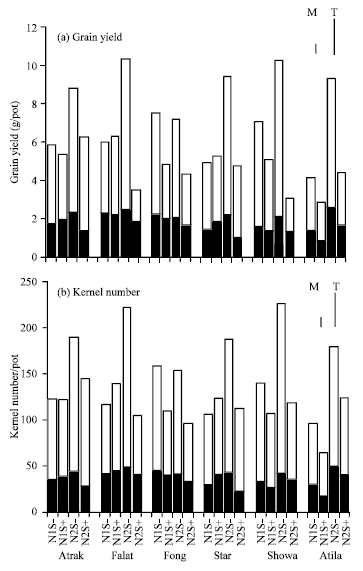 | |
| Fig. 4: | Grain yield and kernel numbers from the main stem (■) and the tillers (□) of six cultivars growth at 2 N rates without post-anthesis water stress (S-) or with post-anthesis water stress (S+). Vertical lines are the LSD (p = 0.05) for the main stem (M) and tillers (T) |
The number of kernel per plant in Star was reduced by stress at the high N rate (Table 2), mainly because of the reduction of kernel in the tillers, but kernel weight of grain from the tillers was less affected by stress than that of Falat, Showa and Atila. Fong showed no response to N in either the main stem or the tillers and stress reduced the yield similarly in both plant parts.
Kernel weight of grain from the tillers was less than that from the main stem and with the exception of Atrak, the variation between treatments was greater (Fig. 5).
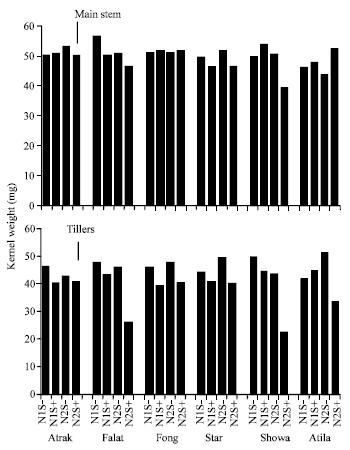 | |
| Fig. 5: | Kernel weight from the main stem and the tillers of six wheat cultivars growth at 2 N rates without post-anthesis water stress (S-) or with post-anthesis water stress (S+). Vertical lines are the LSD (p = 0.05) for the main stem and tillers |
Kernel weight was reduced by stress, particularly in the high n treatment. The main stem produced grain with a higher GPC than the tillers on average and the GPC was more responsive to N than grain the tillers (Fig. 6).
DISCUSSION
Significant differences in the responses to N and water stress were measured between cultivars. Large increase in grain yield occurred in Falat and Atila, with Atila being the most responsive to N, whereas Fong did not respond to N. These were also difference numbers of kernels set. There were also differences among the cultivars in the sensitivity of kernel weight to high N and post-anthesis stress, which also reflected yield responses.
The level of N significantly affected the response to stress. Post-anthesis stress at the low N rate did not significantly reduce grain yield and did not substantially affect kernel weight and GPC, but when post-anthesis stress was imposed on plants grown at high N rate, GPC was increased significantly and grain yield and kernel weight were reduced.
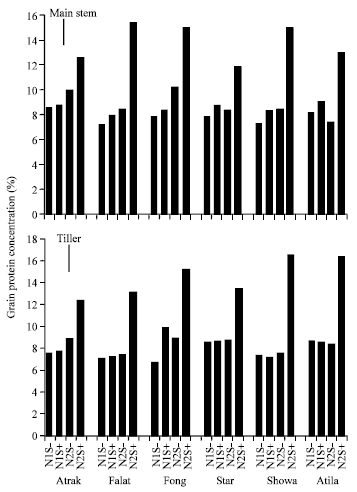 | |
| Fig. 6: | Grain protein concentration from the main stem and the tillers of 6 wheat cultivars grown at 2 N rates without post-anthesis water stress (S-) or with post-anthesis water stress (S+). Vertical lines are the LSD (p = 0.05) for the main stem and tillers |
The response was particularly evident in Atila, Falat and Showa, where yield and kernel weights of stressed plants at the high N level were lower than of stressed plants at the low N level. This perhaps, illustrates the rick associated with applying N to wheat in environments where post-anthesis drought occurs. The varietal differences in the response suggest that wheat production based on sensitive cultivars such as Falat and Atila needs to be confined to regions where the probability of post-anthesis moisture stress is low.
The differences in the response to N and water stress among the genotypes were associated with the responses of the tillers. The main stem contributed 22-54% of the total yield and although N and stress had similar effects on the yield of the main stem and tillers, the responses were much greater in the tillers. Turner et al. (1994) contend that the progressive development of post-anthesis water stress results in late-formed tillers being exposed to higher levels of stress during flowering and grain filling, which reduces their yield. In this experiment, there also appear to be important genotypic differences in the responses to stress associated with differences in the rates of tiller development within the plant but which do not seem to be directly related to tiller number per se. At the higher level of N, tiller and ear numbers were increased in Falat and Showa but stress greatly reduced Kernel number and kernel weight in the tillers. However, in Atila, which produced a greater number of tillers and ears than Falat or Showa, kernel numbers and kernel weight were not reduced as much. The high proportion of fertile tillers observed at the first harvest suggests that Atila had a more synchronous tillering habit than Falat or Showa. In these latter cultivars, the more extended time over which ear emergence occurred may have exposed the later formed ears to greater levels of stress than the ears of Atila, inducing larger reductions in grain set and kernel growth. Differences in the rates of tiller and ear development among genotypes may have affected yields of the tillers and their relative sensitivity to stress by exposing the late formed tillers to higher level of stress and altering the competitive relationships between the main stem and the tillers. Consequently, the contribution of tillers to grain yield was greatly reduced (Table 4). However, differences in tillering do not explain all of the genotypic differences in yield sensitivity.
Genotypic differences in tolerance to stress not related to tillereing were evident in the experiment. Atrak showed less response to the N and stress treatments than Falat and Showa, although they had similar numbers of tillers and a similar proportion of fertile tillers at H1. Brook et al. (1982) found that water stress reduced the kernel weight of Atrak by only 3-5 mg compared with 10 mg in wheat. These observations suggest that Atrak is s cultivar that a relatively stable kernel weights. Star, which is related to Atrak, also showed a relatively stable kernel weight in the tillers, but unlike Atrak, kernel numbers in the tillers were reduced by stress. In this case, the maintenance of kernel of weight may have been associated with the reduction in kernel numbers. If there is not a large reduction in kernel numbers per ear which alters the sink to source ratio, the 2 possible ways by which kernel growth and final kernel size can be maintained under stress are by maintaining a supply of photoassimilate to the ear from concurrent photosynthesis and remobilization (maintaining the source) and by the grain having the ability to continue starch deposition as stress intensifies (maintaining sink strength). The data from the present experiment showed that net remobilization increased with stress and that there was genotypic difference in the net amount of dry matter remobilised, but this was not directly associated with the stability in kernel weight. Therefore, the latter mechanism (maintaining sink strength) may be more likely.
Studies using a single cultivars have shown that remobilization of dry matter and its contribution to grain yields increases with water stress (Bidinger et al., 1977; Reuter and Robinson, 1985; Palta et al., 1994). Estimates in the present experiment based on the change in dry matter indicates considerable variation in the amounts of dry matter remobilised, although our measurements of dry matter changes are unable to distinguish between respiratory losses and the amount of dry matter translated to the grain. The proportion of the total amount remobilised which is respired may not be affected as much by stress as the total amount respired (Turner et al., 1994; Palta and Fillery, 1995b) and we assumed that the net dry matter renobilised estimated in this experiment reflected the relative differences in the dry matter translocated to the grain, if not the actual amount. Despite measuring genotypic differences in dry matter remobilization there was no association between the amount of dry matter remobilised by a cultivar and the stability of kernel weight between the treatments. For example, the kernel weight of Falat, a cultivar in which net remobilization was high, was very sensitive to post-anthesis stress, whereas Atrak, which had a much less variable kernel weight, remobilised less dry matter. Other studies in wheat have also found little relationship between the ability to remobilised dry matter and kernel weight stability among genotypes (Hossain et al., 1990; Savin and Nicolas, 1964), whereas data of Siddique et al. (1989) indicated an inverse relationship between the average kernel weight and the proportion of dry matter remobilised at anthesis that is, low remobilization was associated with heavy kernels. In a comparison of 2 cultivars of wheat, Blum et al. (1994) found that the cultivar that was able to maintain its kernel weight under stress also had greater stem reserves and remobilised a larger amount of stem dry matter than a cultivar whose kernel weight was sensitive to temperature stress. However, the green leaf area of the former cultivar fell very rapidly, reducing the photosynthetic area and the number of kernels per ear was also greatly reduced at the higher temperatures, both of which affect a simple interpretation of the remobilization results. Savin and Nicolas (1964) found that the loss of stem dry matter decreased by similar amounts in 2 cultivars of wheat under stress, although the kernel weights of the 2 genotypes showed different sensitivities to stress. They concluded that the main effects of post-anthesis stress on the kernel weight of wheat from reduced demond for assimilate by the grain. Although it is clear that there can be considerable remobilization of dry matter to the grain, which contributes to grain growth under post-anthesis stress, these results raise the question of whether there is causal relationship between remobilization of dry matter and kernel weight stability among genotypes or whether it reflects the fact that greater levels of stress result in reductions in grain growth and increases in remobilization independently. Without being able to partition the remobilised dry matter in respiratory loss and that translocated to the grain or the roots, we are unable to answer this, but there may not be a simple relationship because there was no clear association between the net remobilization of dry matter and kernel weight stability among these 6 genotypes. An alternative explanation is that the differences in the sensitivity of kernel weight reflect effects of stress on the processes of starch deposition within the grain. Jenner et al. (1991) considered that the biochemical processes involved in the deposition of starch are sensitive to stress and that genotypic differences in wheat are due to differences in the sensitivity of these intrinsic biochemical pathways. There did appear to be differences in the stability of kernel weight in our experiment: kernel weight were greatly reduced by stress in Atila and Falat and it was notable that the kernel weight of Atrak and Star, which are related cultivars, were relatively more stable under stress than a number of the other cultivars in the experiment. These genotypic differences warrant further investigation. Many of the differences in GPC were related to the effects on kernel weight: GPC increased with post-anthesis stress but the effects were greater at the higher level of N, which replicated the response in kernel weight. Within the plant, the partitioning of N to the grain was not increased by stress, because NHI decreased as results of the lower grain yield. There was a large increase in the GPC of grain from the main stem, owing to water stress at the high level of N, even when there was little change in the mean kernel weight, largely because of an increase in the amount of N per kernel. This indicates that where there is a large pool of N relative to the number of grains set, there can be an increase in the transfer of N to the grain under stress. However, the tillers produced grain with a lower GPC despite setting fewer grain of a lower kernel weight and the increase in N per kernel in the main stem due to stress was greater than that measured in the tillers (25% increase v. 12% increase). These observations suggest that there may have been a preferential movement of N from the tillers to the main stem. This idea is supported by the amounts of N remobilised from the main stem and tillers, which showed that much more of the total plant N was remobilised from the tillers than from the main stem (Fig. 1-3).
The experiment has demonstrated that post-anthesis stress may have greater effects on the yield and GPC of wheat grown under high rates of N. Considerable genotypic variation in the sensitivity to post-anthesis stress was also demonstrated. Yield and GPC of the main stem the tillers were affected by N and stress in a similar manner but part of the variation between genotypes can be attributed to the responses of the tillers and the partitioning of dry matter and N between the tillers and the main stem. The data from the experiment suggest that cultivars that produce a large number of tillers or in which there is a range in the rates of development of tillers will be more susceptible to post-anthesis stress, especially when high rates of N are used that promote tillering. However, differences in tillering do not explain the full range of responses and there appear to be cultivars in which grain weight and grain yield are relatively stable and unrelated to tillering or to the apparent remobilization of dry matter and may be associated with inherent properties of the grain.
REFERENCES
- Birch, C.J. and K.E. Long, 1990. Effect of nitrogen on the growth, yield and grain protein content of barley (Hordeum uulgare). Aust. J. Exp. Agric., 30: 237-242.
CrossRefDirect Link - Brooks, A., C.F. Jenner and D. Aspinall, 1982. Effects of water deficit on endosperm starch granules and on grain physiology of wheat and barley. Aust. J. Plant Physiol., 9: 423-436.
CrossRefDirect Link - El-Alaoui, A.C., S.R. Simmnos and R.K. Crookston, 1992. Allocation of photoassimilate by main shoots and nonsurviving tillers in barley. Crop Sci., 32: 1233-1237.
Direct Link - Hossain, A.B.S., R.G. Sears, T.S. Cox and G.M. Paulsen, 1990. Desiccation tolerance and its relationship to assimilate partitioning in winter wheat. Crop Sci., 30: 622-627.
Direct Link - Kobata, T., J.N. Palta and N.C. Turner, 1992. Rate of development of post-anthesis water deficits and grain filling in spring wheat. Crop Sci., 32: 1238-1242.
Direct Link - Morgan, J.A., 1986. The effects of N nutrition on the water relation and gas exchange characteristics of wheat (Triticum aestivum L.). Plant Physiol., 80: 52-58.
Direct Link - Palta, J.A., T. Kobata, N.C. Turner and I.R. Fillery, 1994. Remobilisation of carbon and nitrogen in wheat as influenced by postanthesis water deficits. Crop Sci., 34: 118-124.
Direct Link - Palta, J.A. and I.R.P. Fillery, 1995. N application increases pre-anthesis contribution of dry matter to grain yield in wheat grown on a duplex soil. Aust. J. Agric. Res., 46: 507-518.
CrossRefDirect Link - Papakosta, K.D. and A.A. Gagianas, 1991. Nitrogen and dry matter accumulation, remobilization and losses for mediterranean wheat during grain filling. Agron. J., 83: 864-870.
CrossRefDirect Link - Pheloung, P.C. and K.H.M. Siddique, 1991. Contribution of stem dry matter to grain yield in wheat cultivars. Aust. J. Plant Physiol., 18: 53-64.
Direct Link - Savin, R.B. and M.E. Nicolas, 1996. Effects of short periods of drought and high temperature on grain growth and strach accumulation of two malting barley cultivar. Aust. J. Plant Physiol., 23: 201-210.
Direct Link - Sofield, I., L.T. Evans, M.G. Cook and I.F. Wardlaw, 1977. Factors influencing the rate and duration of grain filling in wheat. Aust. J. Plant Physiol., 4: 785-797.
CrossRefDirect Link - Turner, N.C., P. Prasertsak and T.L. Setter, 1994. Plant spacing density and yield of wheat subjected to post-anthesis water deficits. Crop Sci., 34: 741-748.
Direct Link - Van Keulen, H. and W. Stol, 1991. Quantitative aspects of nitrogen nutrition in crops. Fertil. Res., 27: 152-160.
CrossRefDirect Link - Zadoks, J.C., T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Res., 14: 415-421.
CrossRefDirect Link - Ehdaie, B. and J.G. Waines, 2001. Sowing date and nitrogen rate effects on dry matter and nitrogen partitioning in bread and durum wheat. Field Crops Res., 73: 47-61.
CrossRef - Bidinger, F.R., R.B. Musgrav and R.A. Fischer, 1977. Contribution of stored pre-anthesis assimilates to grain yield in wheat and barley. Nature, 270: 431-433.
CrossRefDirect Link - Blum, A., B. Sinmena, J. Mayer, G. Golan and L. Shpiler, 1994. Stem reserve mobilisation supports wheat-grain filling under heat stress. Aust. J. Plant Physiol., 21: 771-781.
CrossRefDirect Link