
J.I. Lai
Laboratory of Marine Biotechnology, Institute of Bioscience, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
F.M. Yusoff
Laboratory of Marine Biotechnology, Institute of Bioscience, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
M. Shariff
Laboratory of Marine Biotechnology, Institute of Bioscience, Universiti Putra Malaysia, 43400, Serdang, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 13 | Page No.: 635-640







ABSTRACT
Outdoor mass culture of microalgae in the tropical area is important to minimize its production cost. This study evaluates the growth of Chaetoceros calcitrans in 120 L annular photobioreactors at indoor temperature (Treatment I, 25±2°C) and outdoor tropical ambient temperature, (Treatment II, 30±6°C). Each treatment was done in duplicates. For both treatments, C. calcitrans was first grown in starter columns of 10 L capacity for a period of 7 days at 25±2°C. After 7 days, the 9 L culture was transferred to the annular photobioreactors and subsequently brought to a final volume of 100 L by adding 20 L fresh medium every 5 days. There was no significant difference (p>0.05) in the dry weight of microalgae grown in natural light and those grown indoor. The results suggest that C. calcitrans can be grown in outdoor conditions, hence, saving time and microalgae production cost for the larviculture industry.
PDF Abstract XML References Citation
Received: September 19, 2012;
Accepted: October 19, 2012;
Published: January 23, 2013
How to cite this article
J.I. Lai, F.M. Yusoff and M. Shariff, 2012. Large-scale Culture of a Tropical Marine Microalga Chaetoceros calcitrans (Paulsen) Takano 1968 at Different Temperatures Using Annular Photobioreactors. Pakistan Journal of Biological Sciences, 15: 635-640.
DOI: 10.3923/pjbs.2012.635.640
URL: https://scialert.net/abstract/?doi=pjbs.2012.635.640
DOI: 10.3923/pjbs.2012.635.640
URL: https://scialert.net/abstract/?doi=pjbs.2012.635.640
INTRODUCTION
Microalgae form a very important component of the live feed production system in aquaculture hatcheries due to their high content of valuable biochemical products such as antioxidants and polyunsaturated fatty acids (Natrah et al., 2007; Goh et al., 2009, 2010). Essential live feed organisms such as rotifers and copepods require microalgae not only as feed items, but also for enrichment of their nutritive values (Farhadian et al., 2008, 2009; Khatoon et al., 2009; Banerjee et al., 2011; Khatoon et al., 2012). Due to their important role in providing a feed resource in aquaculture production, simple large scale microalgae production is necessary for larviculture industry. In the past, most hatcheries in Asia have been using plastic bags, tanks and open ponds for their algae culture. Our preliminary studies indicate that the production of the microalgae in plastics bags is about ten times lower than that propagated in photobioreactors. Efficient production of microalgae in photobioreactors are now necessary not only due to rapid development of aquaculture industry but also for other uses such as production of biofuel (Dayananda et al., 2007; Chisti and Yan, 2011; Demirbas, 2011), foods, pharmaceutical and nutraceutical products (Ghasemi et al., 2007; Abd El-Baky et al., 2008).
With more than 90% (62,423,187 tonnes) of world’s aquaculture producers located in Asia (FAO, 2010), a tremendous amount of food is needed to sustain the growing demand of the aquaculture industry. Mass production of microalgae economically without compromising the purity is urgently needed. Most open ponds or tank grown microalgae are at a disadvantage with lower productivity and prone to contaminations (Chisti, 2007; Chen et al., 2009). In addition, outdoor temperatures can be a limiting factor to the growth of microalgae (Mata et al., 2010). Temperature also affects the growth rate of microalgae (Sriharan et al., 1991; Chen et al., 2012; Fukao et al., 2012). Another study conducted by Cho et al. (2007) suggested that temperature has a more significant effect on the maximum density of Nannochloris oculata as compared to salinity. Weiss et al. (1985) reported that although the optimal growth for Tetraselmis suecica was found to be at 25-27°C, cultures were able to tolerate a wider range of temperature (15-32°C), when nutrient supply was abundant. On the other hand, cell metabolism such as nitrate metabolism in Thalassiosira pseudonana decreased when the alga was cultured at suboptimal temperature (Berges et al., 2002).
Under hatchery conditions, environmental factors such as temperature and light can undergo drastic fluctuations which may affect the quality of the microalgae (Veron et al., 1996; Ak et al., 2008). Thus, culture optimization of microalgae species by growing them under ambient temperature is one important strategy for successful and economical larviculture activity. This study was designed to elucidate the capacity of an important live feed species, Chaetoceros calcitrans in 120 L capacity annular photobioreactors under ambient conditions compared to the indoor temperature under which it is normally cultured.
MATERIALS AND METHODS
Microalgae culture: Pure isolate of C. calcitrans (UPMC-A0010) was obtained from Aquatic Animal Health Unit (AAHU), Faculty of Veterinary Medicine, Universiti Putra Malaysia. Natural seawater (30 ppt) filtered through membrane filter paper of 0.45 μm pore size (Sartorius, Germany) was used for preparation of f-medium (Guillard and Ryther, 1962) to grow stock and primary cultures of C. calcitrans. The pH level was adjusted to 7.8-8.2 before autoclaving at 121°C for 15 min.
Experimental design: Four annular photobioreactors (Series F and M-TI 50/40, Zittelli et al., 2006) previously disinfected with sodium hypochlorite and rinsed using filtered tap water through 0.1 μm filter bags, were used in the experiment. Two bioreactors were placed in a temperature (25±2°C) controlled room (Treatment I) while another two were placed under ambient temperature of 30±6°C outdoor under shade (Treatment II). Each bioreactor was inoculated with C. calcitrans at the initial cell concentrations of approximately 4.37x106 cells mL-1 (Treatment I) and 4.36x106 cells mL-1 (Treatment II) in 20 L culture media, using inocula prepared in starter columns. Thereafter, the cultures were diluted with 20 L of fresh medium every 5 days (5th, 10th, 15th, 20th day) until a final volume of 100 L was reached. The cultures were illuminated using two cool daylight fluorescent lamps (cool daylight lamps, OSRAM L58W/865, Germany) set at 12L:12D cycle throughout the experimental period.
Inoculum: Prior to the start of the experiment, four starter columns (10 L capacity) were inoculated with C. calcitrans starting from 1.5 L and were scaled up to 3, 6 and 9 L every 2 days for 7 days. All cultures in the starter columns were grown at 25±2°C and were supplied with air filtered through 0.45 μm air filter (Sartorius, Germany). Industrial pure carbon dioxide was provided to the cultures to maintain the pH at 7.8-8.2 in the culture vessels. The cultures were illuminated using cool daylight fluorescent lamps (cool daylight lamps, OSRAM L58W/865, Germany) set at 12L:12D cycle.
Analysis of growth parameters: Samples were collected daily in the morning (0830-0900) for dry weight measurements. Dry weight of the biomass was determined by filtering a known volume of the cells through a precombusted Whatman GF/C filter paper (105°C, 4 h, cooled and weighed to constant weight) and washed with 0.5 M ammonium formate. The filter paper with biomass was then dried at 105°C for 4 h, cooled and weighed until constant dry weight (modified from Coutteau, 1996). Specific growth rate, μ was estimated using the following formula:
where, W0 is the dry weight at the beginning of the selected time interval (days); Wt is dry weight at the end of the selected time interval (days) and tt-t0 is the time interval (days) for the incubation time.
Statistical analysis: Collected data were analyzed using independent sample t-test. Significant differences were determined at 0.05 level of probability. Statistical analysis was done using SPSS 17.0 for Windows (SPSS Inc. Chicago. IL, USA).
RESULTS
This study illustrated that a diatom, Chaetoceros calcitrans can be grown at outdoor ambient temperature using photobioreactors, indicating that this species could tolerate the daily fluctuations of the local temperature. In fact, cultures grown in outdoor ambient temperature were not significantly different (p>0.05) from those grown indoor.
The average room temperature in Treatment I was 25±2°C while in Treatment II the average room temperature was 30±6°C. The pH ranges for Treatment I and II were 8.6-9.8 and 8.6-9.9, respectively (Table 1). Specific growth rates for Treatment I (0.06/day) and II (0.05/day) were not significantly different (p>0.05).
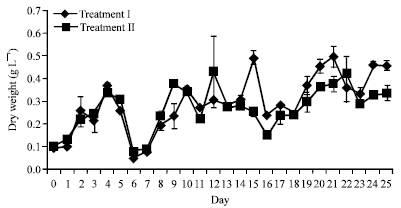 | |
| Fig. 1: | Growth of Chaetoceros calcitrans in 120 L annular photobioreactor measured in terms of dry weight (g L-1), bars represent a 20 L dilution with medium (Mean±SE, n = 2) |
| Table 1: | Ranges of pH of culture, means of culture room temperature and specific growth rates of Chaetoceros calcitrans during the culture period |
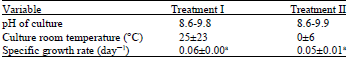 | |
| Values (Mean±SE) in a row with the same superscript are not significantly different at p>0.05 | |
| Table 2: | Means of dry weight in Treatments I and II |
| Value (Mean±SE, n = 2) in each column with same superscript are not significantly different at p>0.05 | |
Upon the first dilution, there was a sharp reduction in biomass in terms of dry weight. Dry weight of C. calcitrans decreased after each dilution (5th, 10th and 15th day) except for the dilution on day 20. The dilution at day 5 reduced the algal concentration to lower than that of the initial concentration (Fig. 1). However, the cultures were able to adapt to this dilution and increased in cell concentration from day 7 onwards.
Generally, the outdoor cultures showed a better growth at the beginning of the study but after day 14, biomass production of indoor cultures surpassed the outdoor cultures. However, a similar trend of biomass production was observed in both indoor and outdoor cultures (Fig. 1). There was no significant difference (p>0.05) between the average dry weight measurements of Treatment I (0.29±0.02 g L-1) and Treatment II (0.28±0.01 g L-1) (Table 2). Maximum biomass achieved were (0.49 g L-1) and (0.39 g L-1) for indoor and outdoor cultures, respectively.
DISCUSSION
In the present study of C. calcitrans large scale culture, there were no significant differences in the growth rates in Treatment I and Treatment II, indicating that the microalgae could quickly adapt to the new temperature without prior acclimatization (Fig. 1). Chaetoceros calcitrans was originally grown indoor (25±2°C) and our results showed that C. calcitrans was able to tolerate the high ambient temperature range of 24-36°C. Renaud et al. (2002) demonstrated that Chaetoceros sp. grew best at 30°C but also showed that tolerance to high temperatures of 33 and 35°C with a moderate growth rate of 0.53/day. This finding was further supported by Araujo and Garcia (2005) and Banerjee et al. (2011) who reported that C. calcitrans grow best at temperature ranging from 25-36°C. The continuous growth of C. calcitrans after every dilution is an indication that it was able to adapt to the sudden changes in environmental conditions (Fig. 1). During the experiment, three growth phases were observed. Lag phase of 7 days was observed while exponential phase was observed thereafter. The cultures progressed into stationary phase after day 10 as the growth rate was reduced. The sharp decrease after the first culture medium addition could be due to the dilution while the cultures were still in adaptation stage and the cell density was still low. Nevertheless, cultures in both treatments managed to adapt to their respective culture environments. Probably, the addition of fresh culture medium to the bioreactor provided adequate nutrients to the microalgae cells (Fig. 1).
In the natural environment, the effect of temperature is not only shown physically by changes in growth but also at cell metabolism level such as photosynthesis, carbon and nitrogen uptake (Anning et al., 2001; Berges et al., 2002; Araujo and Garcia, 2005). Anning et al. (2001) demonstrated that light harvesting pigments increased in abundance with increasing temperature and suggested that at low temperatures, carbon fixation mediated by photosynthetic enzymes is affected. Meanwhile, nitrogen uptake in Thalassiosira pseudonana was shown to decrease with increasing temperatures suggesting that nitrate metabolism in this diatom is mediated through enzymatic activities (Berges et al., 2002). In fact, other factors could also modulate the effects of temperature on microalgae growth. For instance, Chinnasamy et al. (2009) reported that high temperature (50°C) reduced the growth of Chlorella vulgaris cultures when 6% of CO2 was provided to the cultures and those grown at ambient CO2 levels showed no growth. Addition of carbon dioxide is also reported to alter the biochemical composition of microalgae through a higher production of protein Araujo and Garcia (2005) while extending the exponential phase of algal culture (Fabregas et al., 2001).
In the present study, there was no significant difference (p>0.05) in the biomass production of cultures maintained at temperature 25±2°C (treatment I) and ambient temperature of 30±6°C (treatment II). Average specific growth rate was 0.06/day and 0.05/day for treatment I and II, respectively. McGinnis et al. (1997) demonstrated that Chaetoceros muelleri achieved optimum growth rate at 30°C with 4.0 doublings/day as compared to those cultured at 20°C while Banerjee et al. (2011) also reported that C. calcitrans grew better in outdoor conditions as compared to laboratory cultures in 1 L culture medium. However, cultures in the previous studies were not grown in a large volume as in the present study where 120 L photobioreactors were used. Apart from that, a previous study by Banerjee et al. (2011) also pointed out that the cultures were acclimatized to the study conditions, whereas in the present study, inoculums were grown indoor before the study period. Thus, further studies on the acclimatization of microalgae to outdoor conditions and biomass production in annular photobioreactors should be carried out to elucidate the effects of temperature on microalgae.
Culturing microalgae in outdoor environment can achieve higher yields, leading to a reduction of operating cost compared to cultures cultivated indoors (Lopez-Elias et al., 2005). In the present study, biomass production in terms of dry weight in treatment II at the end of the study (25 days, 0.35 g L-1) was similar to the biomass achieved during the first 10 days of the experiment (0.36 g L-1). It can be suggested that microalgae cultured in treatment II conditions can be harvested earlier. The biomass obtained in the present study was lower than the previous study in 1 L (Banerjee et al., 2011), probably due to the fact that the 20 L dilution was done every 5th day.
CONCLUSION
Chaetoceros calcitrans can be mass cultivated in the outdoor tropical ambient temperatures. This study suggested that cultivation of C. calcitrans under ambient temperature conditions using photobioreactors were able to produce higher biomass in a shorter period of time. In fact, there was no significant difference between the outdoor and indoor mass culture (p>0.05). Further studies are needed to establish standard protocol for large scale production of microalgae in photobioreactors.
ACKNOWLEDGMENTS
We would like to thank Dr. Hazel Monica Matias-Peralta for isolating the C. calcitrans used in this study. We would also like to thank Dr. Sanjoy Banerjee, Dr. Helena Khatoon and Mr. Perumal Kuppan for their technical assistance. This study was funded by Johor Satellite Biotechnology Project grant No. BSP(J)/BTK001(4).
REFERENCES
- Abd El-Baky, H.H., F.K. El Baz and G.S. El-Baroty, 2008. Characterization of nutraceutical compounds in blue green alga Spirulina maxima. J. Med. Plants Res., 2: 292-300.
Direct Link - Ak, I., S. Cirik and T. Goksan, 2008. Effects of light intensity, salinity and temperature on growth in camalti strain of Dunaliella viridis teodoresco from Turkey. J. Boil. Sci., 8: 1356-1359.
CrossRefDirect Link - Anning, T., G. Harris and R.J. Geider, 2001. Thermal acclimation in the marine diatom Chaetoceros calcitrans (Bacillariophyceae). Eur. J. Phycol., 36: 233-241.
CrossRef - Banerjee, S., W.E. Hew, H. Khatoon, M. Shariff and F.M. Yusoff, 2011. Growth and proximate composition of tropical marine Chaetoceros calcitrans and Nannochloropsis oculata cultured outdoors and under laboratory conditions. Afr. J. Biotechnol., 10: 1375-1383.
Direct Link - Berges, J.A., E.V. Diana and P.J. Harrison, 2002. Effects of temperature on growth rate, cell composition and nitrogen metabolism in the marine diatom Thalassiosira pseudonana (Bacillariophyceae). Mar. Ecol. Prog. Ser., 225: 139-146.
Direct Link - Chen, P., M. Min, Y. Chen, L. Wang and Y. Li et al., 2009. Review of the biological and engineering aspects of algae to fuels approach. Int. J. Agric. Biol. Eng., 2: 1-30.
Direct Link - Chen, S.Y., L.Y. Pan, M.J. Hong and A.C. Lee, 2012. The effects of temperature on the growth of and ammonia uptake by marine microalgae. Bot. Stud., 53: 125-133.
Direct Link - Zittelli, G.C., L. Rodolfi, N. Biondi and M.R. Tredici, 2006. Productivity and photosynthetic efficiency of outdoor cultures of Tetraselmis suecica in annular columns. Aquaculture, 261: 932-943.
CrossRef - Chinnasamy, S., B. Ramakrishnan, A. Bhatnagar and K.C. Das, 2009. Biomass production potential of a wastewater alga Chlorella vulgaris ARC 1 under elevated levels of CO2 and temperature. Int. J. Mol. Sci., 10: 518-532.
CrossRefDirect Link - Chisti, Y. and J. Yan, 2011. Energy from algae: Current status and future trends: Algal biofuels-a status report. Applied Energy, 88: 3277-3279.
CrossRef - Cho, S.H., S.C. Ji, S.B. Hur, J. Bae, I.S. Park and Y.C. Song, 2007. Optimum temperature and salinity conditions for growth of green algae Chlorella ellipsoidea and Nannochloris oculata. Fish. Sci., 73: 1050-1056.
CrossRefDirect Link - Dayananda, C., R. Sarada, M. Usha Rani, T.R. Shamala and G.A. Ravishankar, 2007. Autotrophic cultivation of Botryococcus braunii for the production of hydrocarbons and exopolysaccharides in various media. Biomass Bioenergy, 31: 87-93.
CrossRefDirect Link - de Castro Araújo, S. and V.M.T. Garcia, 2005. Growth and biochemical composition of the diatom Chaetoceros cf. wighamii brightwell under different temperature, salinity and carbon dioxide levels. I. Protein, carbohydrates and lipids. Aquaculture, 246: 405-412.
CrossRefDirect Link - Fabregas, J., A. Otero, A. Dominguez and M. Patino, 2001. Growth rate of the microalga Tetraselmis suecica changes the biochemical composition of Artemia species. Mar. Biotechnol., 3: 256-263.
CrossRefDirect Link - Demirbas, M.F., 2011. Biofuels from algae for sustainable development. Applied Energy, 88: 3473-3480.
CrossRef - Farhadian, O., F.M. Yusoff and A. Arshad, 2008. Population growth and production of Apocyclops dengizicus (Copepoda: Cyclopoida) fed on different diets. J. World Aquacult. Soc., 39: 384-396.
CrossRef - Farhadian, O., F.M. Yusoff and S. Mohamed, 2009. Nutritional values of Apocyclops dengizicus (Copepoda: Cyclopoida) fed Chaetocerous calcitrans and Tetraselmis tetrathele. Aquacult. Res., 40: 74-82.
CrossRef - Fukao, T., K. Kimoto and Y. Kotani, 2012. Effect of temperature on cell growth and production of transparent exopolymer particles by the diatom Coscinodiscus granii isolated from marine mucilage. J. Applied Phycol., 24: 181-186.
CrossRef - Ghasemi, Y., A. Moradian, A. Mohagheghzadeh, S. Shokravi and M.H. Morowvat, 2007. Antifungal and antibacterial activity of the microalgae collected from paddy fields of Iran: Characterization of antimicrobial activity of Chroococcus dispersus. J. Boil. Sci., 7: 904-910.
CrossRefDirect Link - Goh, Jr. L.P., S.P. Loh, M.Y. Fatimah and K. Perumal, 2009. Bioaccessibility of carotenoids and tocopherols in marine microalgae, Nannochloropsis sp. and Chaetoceros sp. Mal. J. Nutr., 15: 77-86.
PubMedDirect Link - Goh, S.H., F.M. Yusoff and S.P. Loh, 2010. A comparison of the antioxidant properties and total phenolic content in a diatom, Chaetoceros sp. and a green microalga, Nannochloropsis sp. J. Agric. Sci., 2: 123-130.
CrossRefDirect Link - Guillard, R.R. and J.H. Ryther, 1962. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J. Microbiol., 8: 229-239.
CrossRefDirect Link - Khatoon, H., S. Banerjee, F.M. Yusoff and M. Shariff, 2009. Evaluation of indigenous marine periphytic Amphora, Navicula and Cymbella grown on substrate as feed supplement in Penaeus monodon postlarval hatchery system. Aquacult. Nutr., 15: 186-193.
CrossRef - Khatoon, H., S. Banerjee, F.M. Yusoff and M. Shariff, 2012. Use of microalgal-enriched Diaphanosoma celebensis Stingelin, 1900 for rearing Litopenaeus vannamei (Boone, 1931) postlarvae. Aquacult. Nutr.
CrossRefDirect Link - Lopez-Elias, J.A., D. Voltolina, F. Enriquez-Ocana and G. Gallegos-Simental, 2005. Indoor and outdoor mass production of the diatom Chaetoceros muelleri in a Mexican commercial hatchery. Aquacult. Eng., 33: 181-191.
CrossRef - Mata, M.T., A.A. Martins and N.S. Caetano, 2010. Microalgae for biodiesel production and other applications: A review. Renewable Sustainable Energy Rev., 14: 217-232.
CrossRefDirect Link - McGinnis, K.M., T.A. Dempster and M.R. Sommerfeld, 1997. Characterization of the growth and lipid content of the diatom Chaetoceros muelleri. J. Applied Phycol., 9: 19-24.
CrossRefDirect Link - Natrah, F.M.I., F.M. Yusoff, M. Shariff, F. Abas and N.S. Mariana, 2007. Screening of Malaysian indigenous microalgae for antioxidant properties and nutritional value. J. Applied Phycol., 19: 711-718.
CrossRef - Renaud, S.M., L.V. Thinh, G. Lambrinidis and D.L. Parry, 2002. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures. Aquaculture, 211: 195-214.
CrossRefDirect Link - Sriharan, S., D. Bagga and M. Nawaz, 1991. The effects of nutrients and temperature on biomass, growth, lipid production and fatty acid composition of Cyclotella cryptica Reimann, Lewin and Guillard. Applied Biochem. Biotechnol., 28-29: 317-326.
CrossRef - Veron, B., C. Billard, J.C. Dauguet and M.A. Hartmann, 1996. Sterol composition of Phaeodactylum tricornutum as influenced by growth temperature and light spectral quality. Lipids, 31: 989-994.
CrossRef