
N.T. Ngongoni
Department of Animal Science, University of Zimbabwe, P.O. Box MP 167, Mt Pleasant, Harare, Zimbabwe
C. Mapiye
Department of Livestock and Pasture Sciences, Faculty of Science and Agriculture, University of Fort Hare, Private Bag X1314, Alice 5700, South Africa
M. Mwale
Department of Livestock and Pasture Sciences, Faculty of Science and Agriculture, University of Fort Hare, Private Bag X1314, Alice 5700, South Africa
B. Mupeta
Plan International, P. Bag 7232, Highlands, Harare, Zimbabwe
M. Chimonyo
Department of Livestock and Pasture Sciences, Faculty of Science and Agriculture, University of Fort Hare, Private Bag X1314, Alice 5700, South Africa
Pakistan Journal of Nutrition
Year: 2009 | Volume: 8 | Issue: 4 | Page No.: 377-383







ABSTRACT
Two dairy concentrates (ram press sunflower and sunflower heads) were formulated locally and their effects on the performance of crossbred dairy cows were compared with that of a commercial dairy concentrate. There were no significant differences in body condition and milk production responses by crossbred dairy cows to sunflower based diets compared to commercial dairy concentrates (p > 0.05). It was suggested that the sunflower based rations can be used as cheaper alternatives to conventional dairy concentrates. The ram press sunflower cake, in particular is an option that may be used by smallholder dairy farmers to formulate local dairy concentrate, which influence dairying to a similar extent as the commercial dairy concentrate but cheaply. More research is required to determine fermentation patterns, rumen microbial protein synthesis, true intestinal digestibility and absorption of nutrients from sunflower-based diets at the small intestines in crossbred dairy cows.
PDF Abstract XML References Citation
How to cite this article
N.T. Ngongoni, C. Mapiye, M. Mwale, B. Mupeta and M. Chimonyo, 2009. Sunflower Based Rations for Small-Medium Milk Producing Dairy Cows. Pakistan Journal of Nutrition, 8: 377-383.
DOI: 10.3923/pjn.2009.377.383
URL: https://scialert.net/abstract/?doi=pjn.2009.377.383
DOI: 10.3923/pjn.2009.377.383
URL: https://scialert.net/abstract/?doi=pjn.2009.377.383
INTRODUCTION
Milk production in the smallholder dairy sector is mostly constrained by shortage of affordable appropriate nutritional regimes and overall management. The ever spiraling cost of commercial protein supplements makes it imperative for the smallholder farmers to look for and turn to relatively cheap nitrogen sources for their cows or to be more efficient in their rationing of protein to their low to medium producing crossbred dairy cows (Ngongoni et al., 2007a). Past research indicates that legume forage based rations offered potential to increase milk yield per cow, especially milk protein production (Clatworthy and Holland, 1979). It has been reported, (Orskov, 1970) that for low to medium producing dairy cows, if enough rumen degradable protein is fed to promote maximum rumen microbial protein synthesis, it is likely to meet total protein requirements of that category of lactating dairy cows and other biological demands of the animal.
Sunflower is an alternative protein and energy supplement for farmers located in marginal areas where soybean and maize cannot be grown successfully and the use of whole rather than ground seeds would be easier for on-farm feeding (Ngongoni et al., 2007b). Research has been conducted on optimum levels of inclusion of whole sunflower seed in dairy rations as fat supplements that increase energy density to meet energy requirements of high producing dairy cows in early lactation (Palmquist and Jenkins, 1980; Rafalowski and Park, 1982; Petit, 2003; He et al., 2005). Sunflower seeds increase the proportion of unsaturated fatty acids (especially linoleic acid) in milk fat, without affecting dry matter intake (DMI), milk production or composition (Schingoethe et al., 1996; Petit et al., 2004; He et al., 2005). Sunflower seed (partially dehulled or fully dehulled) produced similar milk yield results to soybean meal in dairy cow rations (Schingoethe, 1981 and Drackley and Schingoethe, 1986). The contribution of rumen microbial protein to total amino acids flowing to the small intestines ranges from 50-90 % (Buttery and Foulds, 1978). With poor quality forage-based diets prevalent in the smallholder dairy sector, microbial protein production in the rumen is often the main source of protein flowing to and is truly digested in the small intestines (Ngongoni et al., 2007b). It, therefore, should ideally be maximized (Perez et al., 1997). Obviously, the recommended feeding strategy for such low cost dairy production systems is to supply adequate rumen degradable protein supplements for low to medium quality basal forage diets to maximize rumen microbial protein synthesis. This would allow optimum degradation of forages and finally increase the supply of truly digestible amino nitrogen to the small intestines, which should meet total protein needs of the low/medium yielding crossbred dairy cows.
The use of the ram press sunflower residue from oil extraction and crushed sunflower heads as a source of protein and energy in dairy concentrates for moderate milk production using crossbred dairy cows (Bos taurus x Bos indicus) has not been studied. Therefore, the objective of this study was to compare conventional commercial dairy concentrate with diets largely basedon ram-press on-farm sunflower residues and crushed sunflower heads as the main sources of nitrogen for low/medium levels of milk production by crossbred smallholder dairy cows.
MATERIALS AND METHODS
Animals: Thirty-six F1 cross Guernsey X Nkone cows bred at Henderson Research Station were used for the current lactation study. The Guernsey heifers conceived to natural mating using Nkone bulls over 90 days bulling period and the heifers calved down within 8 weeks of each other. Their mean body condition score after calving was 3.29 ± 0.16 units and had a mean initial post - partum live - weight of 380 ± 8.4 kg. During pregnancy, the group of cows were grazing natural pastures dominated by Hyperrhenia rodis and Hyperrhenia nyasa species in a three paddock rotational grazing system, at a stocking rate of 3.5 hectares per one livestock unit.
Diets: The treatment rations compared were a commercially manufactured dairy concentrate (control-CDC) an on-farm mixed dairy ration using ram-press-oil-extracted sunflower seed residues (SF) and a third dairy ration formulated using crushed whole sunflower head (excluding seeds) and urea (SF head). The sunflower seed (Panar variety) grown and harvested at Henderson Research Station farm, was pressed using the on-farm ram press machine to extract the oil. The SF treatment diet was produced after proximate analysis of the residues and mixing on-farm using a commercial feed mixer such that sunflower residues comprised 340 kg/tonne(t); crushed maize grain 591 kg/t and a 15 kg/t of Henderson mineral/vitamin premix. The maize grain was crushed through a hammer mill without a sieve and the Hamish Cameron Pvt. Ltd., Harare, supplied the mineral/vitamin premix. For the SF head dairy concentrate, seed was removed from the sunflower heads before they were milled with the same mill as the maize grain. Then the mixture comprised sunflower heads of 408 kg/t; crushed maize grain of 408 kg/t and to raise the crude protein content of the ration, 11 kg/t urea and 164 kg/t of cotton seed cake were added and finally, 15 kg/t of the vitamin and mineral premix. The commercial dairy concentrate used as the control treatment was purchased every three months and samples analyzed to make other two treatment diets i.e SF and SF head, iso-nitrogenous with the control diets using the DAA feed computer programme (Denmark). The actual composition of the three treatment diets CDC, SF and SF head is shown in Table1 with an iso-nitrogenous mean crude protein of 147.0 ± 0.04gkg-1 DM and an energy density of 11.9 ± 0.06 MJ/kg DM.
Experimental design and procedure: The thirty-six heifers were divided into three groups of twelve that were balanced of weight and expected calving date. The groups of heifers were then assigned to each of the treatments in a randomized block design where live-weight and expected dates of calving were used as the blocking factors. Two months before the expected date of calving, all experimental heifers were receiving 2 kg/day of a mixture of cotton seed cake, crushed maize grain and Katambora Rhodes grass hay. The experiment started when the first seven cows calved within three days of each other and after the cows were adapted to the milking routine for one month and as well as to their respective rations for seven days. The cows were machine milked daily (at 0700 and 1500 hours). For 5 days after calving, the calf received colostrums or milk by bottle - drenching after which the cow's milk yield was weighed and recorded. From seven days post-calving, the calf was fostered to a different cow until it was weaned at 60 days. Daily milk samples of the morning and afternoon milking were collected fortnightly until the cows were dried. The cows were weighed within twenty-four hours of calving and at fortnightly intervals thereafter. The cows were weighed using a weigh bridge and body condition scored by the same technician at the same time of the scoring day throughout the trial period.
The cows on trial were given 4.5kg DM of the respective concentrate treatment daily throughout the lactation period and that supplied approximately 662g of crude protein and 54 MJ ME of energy. The rationing was sufficient to meet the requirements for the production of about 10kg of milk (ARC, 1984). Each respective treatment ration was given individually in pens between 07: 00 and 09: 00 h after the morning milking. From the time cows completed their treatment rations, there were grazed together and had equal access to rangeland resources from which it was expected that they would obtain all their maintenance requirements estimated to be 250g of crude protein and 54 MJ ME of energy per day (ARC, 1984). The nutritive quality of natural pastures was monitored during the summer through regular random sampling for proximate analysis from the paddocks on the fifteenth day every month. Depending on the analytical results of the sampled natural pasture, grass silage was given when the crude protein content of the natural pasture declined below 7 %. Grass silage was rationed after individual concentrate allocation refusals were collected and weighed back to assess the concentrates and silage intakes. Borehole water was freely accessible to grazing cows in the paddocks. A spray race was used to spray Diamide acaricide weekly during summer and fortnightly during the dry season. The samples of treatment rations, roughages and refusals were also collected at feeding for proximate or detergent analyses.
Chemical analysis: The DM intake of silage and dairy rations was determined by the difference between dry matter offered and that refused. The Proximate, detergent and chemical analysis were conducted on the samples, after the determination of DM.
| Table 1: | Chemical composition of treatment concentrates and the natural pasture (g/kg of DM) and the daily nutrient intake |
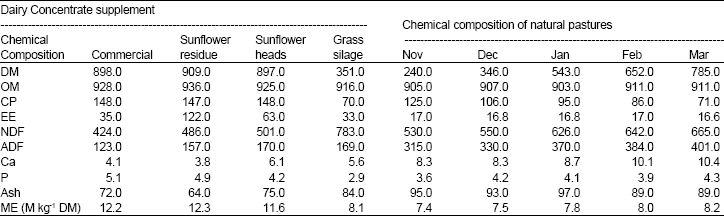 | |
Consequently, dry matter was estimated by drying samples at 105oC for 24 hours and the ash fraction by ashing at 550oC for 12 hours except for the DM determination of silage where a special procedure was adopted to preserve volatile fatty acids. Crude protein, ether attracts, ash and metabolizable energy were determined using the procedures described A.O.A.C. (1990) while Neutral Detergent Fibre (NDF) and Acid Detergent fibre were analyzed using the method of Van Soest et al. (1991). The milk samples were analyzed for butterfat, protein, lactose and total solids using an automated Bentley 2000 Analyzer (Model, B.2000. USA) while A/SN Foss Electronic machine (Model: Fossomatic 90-Denmark) was used to measure somatic cell counts.
Statistical analysis: The data was analyzed using the General Linear Model procedure of the SAS (1987). The model fitted the effects of concentrate type (i.e. CDC, SF and SF head), period of calving (P1 included cows which calved before the on-set of effective rains and P2 included those that calved subsequently) and their interactions on milk yield, milk composition (fat, protein, lactose, total solids and somatic cell counts), post partum live - weight, post partum body condition score and changes in live-weight and body condition through the lactation period. The response to concentrate type was tested over 3 stages of lactation, namely: Early lactation (post partum to 90 days) middle lactation (91 to 231 days post partum) and late lactation (232 to 301 days post partum).
RESULTS
The nutritive composition of the treatment rations is shown on Table 1. Both the dry matter and nutrient intake did not vary considerably with treatment ration except for ether extract whose intake increased significantly (p<0.05) in the SF ration (700 g/day) than SF head (416 g/day) and CDC (293 g/day) rations. In general, all cows on trial cleared all their daily concentrate allocation, with those of the (SF head) treatment diet taking an extra hour to finish their daily allocation.
| Table 2: | Effect of concentrate type lactation phase on yield (kg/day) and composition (g kg-1) of milk from crossbred dairy cows at different stages of lactation |
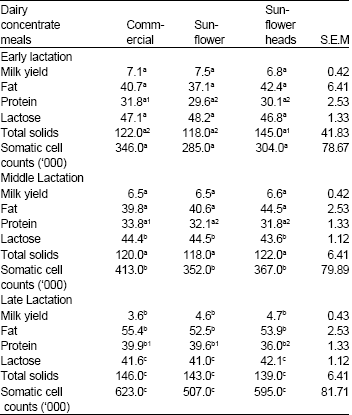 | |
abcMeans of the same nutrient at different stages of lactation in the same column with different superscripts are significantly different (P <0.05). 123Means in the same row with different superscripts are significantly different (P <0.05). | |
Concentrate type had no effect on somatic cells (Table 2). The type of concentrate did not affect the milk yield (p>0.05) during any one of the three stages of lactation (Table 2). However, there was significantly higher (p<0.05) milk yield during early and middle lactation in comparison to late lactation for all treatments. There were no significant differences in milk constituents (butterfat, lactose and total solids) with type of concentrate supplement; except for milk protein, which was high (p<0.05) for CDC. The content of almost all milk constituencies were not significant except lactose, which was higher (p<0.05) during late than early and middle lactation.
| Table 3: | Effect concentrate type and calving period on the performance of crossbred cows. |
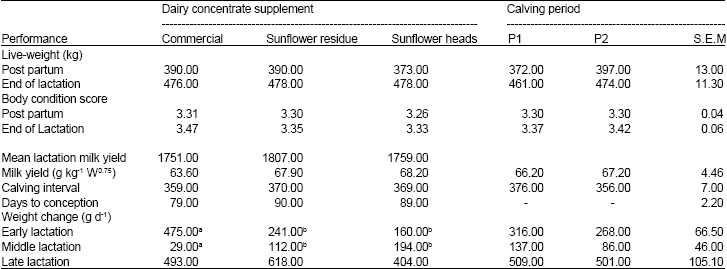 | |
| Means in the same row with different superscripts differ significantly (p<0.05), S.E.M = Standard error of the mean. | |
The cows had a mean lactation length of 296±4 days and mean calving interval of 366±7 days. The type of concentrate had no significant effect on lactation length. Daily milk yield levels reached plateau levels at 9 kg/day for cows given the SF diet and 8.7 kg/day for cows on CDC but for both groups at 10 weeks post partum, while those given the SF head reached plateau levels of 8.2 kg/day at 12 weeks post partum. Milk production declined by 45 % from week 12 to 32 in cows given SF or CDC treatment diets and by 32 % for those given SF head diet. Their overall trend suggest treatment differences in persistence of lactation in that decline in milk production was more marked during late lactation, when milk from cows given CDC declined by 69 %, while those given SF diet declined by 60 % and those given SF head dairy concentrate declined by 54 %.
For all the three concentrate types, there was a gradual decline in the contents of total solids, butterfat, protein and lactose from week 8 to 10 of lactation; reaching the lowest values in week 18 to 20 post partum. Thereafter, total solids and butter fat in milk rose sharply through mid-lactation reaching peak levels during 38 to 42 weeks post partum. During weeks 28 and 38 post partum, cows given the CDC produced milk which had significantly (p<0.05) higher content of total solids and butter fat than milk from cows given the farm produce sunflower concentrates. The protein content of milk in all treatment groups rose steadily from 18 weeks of lactation, giving a sharp rise between 32 through to 42 weeks of lactation. Cows given commercial concentrates maintained significantly higher (p<0.05) protein content throughout the lactation than those receiving farm sunflower-based concentrates. Between 8 and 30 weeks post partum, all milk constituents fluctuated, especially from cows given SF head and CDC than those on the sunflower residue diet. Lactose content in milk from all groups of cows declined through lactation reaching the lowest values at 42 weeks post partum. All milk constituents, between 8 and 30 weeks post partum, tended to fluctuate more in the milk of cows supplemented with SF head or CDC than in milk from SF supplemented cows.
The least squares means for the effect of type of concentrates and calving period on live-weight, body condition, milk production per unit of metabolic body weight and calving interval of crossbred dairy cows are presented in Table 3. At the end of lactation, the type of concentrates had no significant effect (p>0.05) on live-weight, body condition calving interval and efficiency of milk production per unit of metabolic body weight (Table 3). Although there were no significant diet-induced differences in live-weight and body condition, cows given the SF concentrate gained 88kg, while those given CDC gained 86kg and those on SF head gained 75kg during lactation (Table 3). However, concentrate type influenced the rate of live-weight change during the different stages of lactation (p<0.05). The mean daily energy balance and the mean fat-corrected milk yield, milk fat yield and metabolizable energy intake during the 10 weeks of lactation, are shown in Table 4. All cows significantly (p<0.05) lost weight during the first fortnight post-partum. There were significant differences (p < 0.05) in energy balance for sunflower heads (-7.80 MJ/day), sunflower residues (-2.0 MJ/day) and commercial (-1.71 MJ/day) concentrates (Table 4). On average the cows reached energy balance at 14 days post-partum, such that by the fourth week of lactation, the cows were in positive energy balance.
DISCUSSION
Similar to findings of this study He et al. (2005) observed no differences in milk production and composition between dairy cows fed sunflower seeds diet and a control diet. However, the incremental milk yield response to the commercial dairy concentrates feeding was 1.33 kg per kilogram of concentrate; which was less than the response of 1.38 and 1.42 kg milk per kg of sunflower cake and sunflower head based concentrates, respectively. Khalili et al. (1992) reported an incremental milk yield response of 0.57 kg milk per kilogram of concentrate.
| Table 4: | Performance of lactating cross bred cows during the first 10 weeks of lactation |
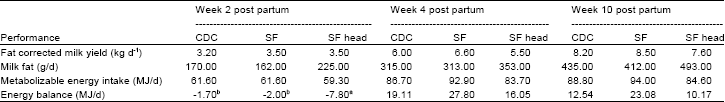 | |
| abMeans with the same superscripts in the same row are not significantly different (P >0.05) | |
Cowan (1971) working with high producing dairy cows, observed a milk yield of 2.2 kg milk per kilogram of concentrate to the feeding of concentrate supplement, which reflects mainly on the genetic merit of cows. The current study shows that it is possible to get a more favourable milk response by crossbred dairy cows to the more cost-effective feeding diets formulated from home-grown sunflower-based diets compared to the relatively expensive convectional commercial dairy concentrates.
Mean daily milk yield and patterns of lactation curves for the three treatment diets indicate higher peak milk yields on CDC and SF treatments than on SF head diet, with no significance in persistence thereafter. In parallel studies on characterization of the three types of concentrates used in the current lactation trial, the effective degradabilities of nitrogen of CDC, SF and SF head treatment diets were, 76.6 %, 88.9 % and 86.3 %, respectively (Ngongoni et al., 2007a). Corresponding values for the amino acids from microbial protein plus UDP truly digested and absorbed in the small intestines [digestibility of the same diets, determined using the mobile nylon bag technique (Hvelplund, 1985), were 76.6 %, 93.5 % and 93.2 %] represent metabolizable protein absorbed and add up to 662, 673 and 560g/kg respectively. Given the above levels of metabolizable protein, to estimate the net efficiency of utilization of absorbed amino nitrogen from milk protein secretion, what is required are estimates of protein requirements for maintenance and the balance goes towards milk protein synthesis. Alternatively, a gross efficiency of nitrogen utilization would be calculated if the metabolizable protein, which gets partitioned towards maintenance is not subtracted. If the values for metabolizable protein supply and subsequent efficiency values are significantly different from each other; it would help to partly explain the lack of significant treatment differences in milk yield. On the basis of the proceeding calculations, the diets used would be expected to elicit 8.9, 9.0 and 6.5 kg milk per day on he CDC, SF and SF head treatments diets, respectively. Interestingly the predicted milk yields are reconcilable with the observed levels of milk production after allowance is made for body weight changes during lactation by the cows on trial.
Cows given the commercial concentrates had significantly higher protein concentration in milk than those given the SF head concentrate. These results provide more evidence that the milk protein concentration in ruminants can be manipulated by dietary means. Moorby et al. (1996) also observed an increase in both milk yield and milk protein content through an increased protein intake by the animals. It was hypothesized that the increase in protein content of milk was probably the consequence of an increase of metabolizable protein supply to post-ruminal sites. The lower concentration of milk protein by cows given the SF head concentrate might be attributed to relatively lower total truly absorbed amino acids in the small intestines on the SF head concentrate than the CDC treatment diet (560 versus 662g/day) (Ngongoni et al., 2007a,b). De Peters and Cant (1992), observed that milk protein content depends on the intake of metabolizable energy (ME) sourced from carbohydrates and that an increase in the fat content of a diet, will tend to reduce protein content. Given the preceding finding and the fact that the daily ME intake was 84.6 MJ ME on SF head diet and 87.8 MJ ME on CDC of which 416 g/day of SF head and 293 g/day of CDC was taken as fat or ether extractives, the low milk protein concentration can be explained by the high intake of fat in the SF head treatment diet.
The observed lactation length is reasonable because very long lactation may lead to increase in calving interval, culminating in low overall cow productivity. Comparable lactation lengths of 270 days were reported for Sahiwal breed (Trail and Gregory, 1982) or 240 days for the Fulani herds in Mali (Wagernaar et al., 1986). In contrast to these data, longer lactation lengths of 470-526 days have been reported for Bunaji and N'Dama cattle in Nigeria and Gambia, respectively (Okyere, 1983; Agyemang et al., 1990) but could be a Bos indicus breed effect. The improvement in lactation yield of cows which calved down after the onset of summer rains reflects an improved nutritional status pre-partum and around calving which probably led to both increased grass growth and subsequent voluntary intake of more nutritious grass. While significant differences in milk production with a range of 2517kg due to period of calving were observed by Trail and Gregory (1981), the milk yield differences between treatments in the current trial did not reach to a significant level probably because the two periods were too close to each other.
The lactation yields reported in the current study are lower than 2000-2500 kg reported for other tropical smallholder dairy cattle breeds such as the Red Sindhi, Sahiwal and Tharparkar used on government cattle research stations (Hodges, 1986). An additional explanation for the lower than expected lactation yield, in the current trial, emanates from the fact that these first calf heifers were mature at calving at ±35 months, these cows produced 75 % of their potential milk production since they were in their first lactation (Trail and Gregory, 1981). Trail and Gregory (1982) reported significant effects of parity on milk shown by below average milk yields for the first and second lactations. Therefore, the observed results are consistent with reports that cattle in the tropics and sub-tropics attain peak production in their third and fourth lactations; which is earlier in the exotic dairy cattle, because cattle in the tropics experience late age at first calving (Mahadevan, 1966).
If the feeding management for lactation is right, live-weight, body condition and conception should generally not be problematic and be strongly related to both nutrient supply and milk production (Butler and Smith, 1989). The normal observed trend is for both milk and feed intake to gradually rise with advancing lactation; but in high producing lactating ruminants, maximum feed intake occurs some weeks after maximum milk production (Schingoethe, 1981; Ngongoni et al., 1989). With reference to live-weight changes in the animal, the consequent negative energy balance that persists for 4 to 12 weeks of lactation is accompanied by mobilization of body tissue reserves to sustain high levels of milk production (Bauman and Currie, 1980; Butler et al., 1981).
Milk production by the cows given CDC increased at the same rate as those on SF diet, both reaching daily yields of 8.7 and 9.0 kg per day at 10 weeks of lactation, respectively. In the current study, crossbred cows remained in negative energy balance for two weeks post partum, which is less than the 4 to 12 weeks post partum (Senatore et al., 1996). Thereafter, all cows gained weight with those on CDC gaining significantly more weight until 12 weeks of lactation and then significantly less weight until 140 days of lactation in comparison to cows on sunflower based diets. The current observation suggest that, although crossbred cows used in this trial went into negative energy balance, the magnitude of body fat mobilization to sustain milk production and the subsequent tissue replenishment does not seem to match that observed in exotic breeds thoroughly bred for high milk production. The causative relationship between milk production, live-weight changes and body tissue mobilization/replenishment and changes in voluntary feed intake in crossbred dairy cows is currently not clearly understood.
Conclusion: The lack of significant treatment differences in milk production responses to sunflower based diets compared to commercial dairy concentrates suggest that the former diets could be used as probable cheaper alternatives to conventional dairy concentrates. The trial indicated that the diets for smallholder dairy cows based on home-grown, ram-pressed sunflower and home-mixed rations can meet the nutritional requirements of crossbred smallholder dairy cows. Because of high fibre content, the SF head diet elicited a higher milk fat content than the SF and CDC diets and lower milk protein content than CDC diet. In addition the SF head diet was less palatable than the ram-press sunflower cake and commercial concentrate based diets. More research is needed on the interaction between live-weight changes, voluntary feed intake, milk yield and composition in crossbred dairy cows. In addition, studies on fermentation patterns, rumen microbial protein synthesis and the flow to post-ruminal sites, true intestinal digestibility and absorption of nutrients from diets at the small intestines are warranted.
ACKNOWLEDGEMENT
The work was financially supported by Danish International Development Agency (DANIDA).
REFERENCES
- Bauman, D.E. and W.B. Currie, 1980. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci., 63: 1514-1529.
CrossRef - Butler, W.R. and R.D. Smith, 1989. Interrelationships between energy balance and postpartum reproductive function in dairy cattle. J. Dairy Sci., 72: 767-783.
CrossRefDirect Link - De Peters, E.J. and J.P. Cant, 1992. Nutritional factors influencing the nitrogen composition of bovine milk: A review. J. Dairy Sci., 75: 2043-2070.
CrossRefPubMedDirect Link - Drackley, J.K. and D.J. Schingoethe, 1986. Extruded blend of soybean meal and sunflower seeds for dairy cattle in early lactation. J. Dairy Sci., 69: 371-384.
CrossRef - Hvelplund, T., 1985. Digestibility of rumen microbial protein and undegraded dietary protein estimated in the small intestine of sheep by in sacco procedure. Acta Agric. Scand., 25: 132-144.
Direct Link - Ngongoni, N.T., C. Mapiye, M. Mwale and B. Mupeta, 2007. Effect of supplementing a high-protein ram press sunflower cake concentrate on smallholder milk production in Zimbabwe. Trop. Anim. Health Prod., 39: 297-307.
CrossRefDirect Link - Palmquist, D.L. and T.C. Jenkins, 1980. Fat in lactating rations: A review. J. Dairy Sci., 63: 11-14.
Direct Link - Petit, H.V., 2003. Digestion, milk production, milk composition and blood composition of dairy cows fed formaldehyde treated flaxseed or sunflower seed. J. Dairy Sci., 86: 2637-2646.
CrossRefDirect Link - Petit, H.V., C. Germiquet and D. Lebel., 2004. Effect of feeding whole, unprocessed sunflower seeds and flaxseed on milk production, milk composition and prostaglandin secretion in dairy cows. J. Dairy Sci., 87: 3889-3898.
Direct Link - Rafalowski, W. and C.S. Park, 1982. Whole sunflower seed as a fat supplement for lactating cows. J. Dairy Sci., 65: 1484-1492.
Direct Link - Schingoethe, D.J., M.J. Brouk, K.D. Lightfield and R.J. Baer, 1996. Lactational responses of dairy cows fed unsaturated fat from extruded soybeans or sunflower seeds. J. Dairy Sci., 79: 1244-1249.
CrossRefPubMedDirect Link - Senatore, E.M., W.R. Butler and P.A. Oltenacu, 1996. Relationships between energy balance and post partum ovarian activity and fertility in first lactation dairy cows. Anim. Sci., 62: 17-23.
Direct Link - Trail, J.C.M. and K.E. Gregory, 1982. Production characters of the Sahiwal and Ayrshire breeds and their crosses in Kenya. Trop. Anim. Health Prod., 14: 45-57.
CrossRefDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link