
I. Batubara
Department of Chemistry, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, IPB Darmaga Campus, Bogor, Indonesia
S. Kotsuka
Faculty of Applied Biological Science, Gifu University, 1-1 Yanagido, Gifu, 501-1193, Japan
K. Yamauchi
Faculty of Applied Biological Science, Gifu University, 1-1 Yanagido, Gifu, 501-1193, Japan
H. Kuspradini
Faculty of Forestry, Mulawarman University, Samarinda, East Kalimantan, Indonesia
T. Mitsunaga
Faculty of Applied Biological Science, Gifu University, 1-1 Yanagido, Gifu, 501-1193, Japan
L.K. Darusman
Department of Chemistry, Faculty of Mathematics and Natural Sciences, Bogor Agricultural University, IPB Darmaga Campus, Bogor, Indonesia
Research Journal of Medicinal Plants
Year: 2012 | Volume: 6 | Issue: 6 | Page No.: 406-415







ABSTRACT
In this study 28 Indonesian medicinal plants species has been screened as potential Tumor Necrosis Factor (TNF)-α inhibitor. Plant materials were extracted with methanol. The activity test was performed on human monocyte-derived THP-1 (ATCC No. TIB 202) which was stimulated by Lipopolyssacharide (LPS). The TNF-α production and cell viability were determined by commercial ELISA kit and WST-1, respectively. All extracts were determined for their phenolic, flavonoid and tannin contents by spectrophotometry and chromatography methods. From 40 plant materials, 5 plant materials had cell viability lower than 70% at 10 ppm concentration and about 8 plant materials had good cell viability higher than 90% at 10 ppm concentration. Among 8 plant materials extract which had good cell viability, only Avicenia sp. stem and Goniothalamus macrophyllus leaf methanol extracts had high TNF-α inhibitory activity. The total phenolic, flavonoid and tannin contents were varied depending on the sample. No correlation was found between TNF-α inhibitory activity with the total phenolic content, flavonoid content and tannin content. In conclusion, among Indonesian medicinal plants, the highest potential as TNF-α inhibitors were Avicenia sp. stem and G. macrophyllus leaves.
PDF Abstract XML References Citation
Received: November 03, 2011;
Accepted: December 29, 2011;
Published: March 15, 2012
How to cite this article
I. Batubara, S. Kotsuka, K. Yamauchi, H. Kuspradini, T. Mitsunaga and L.K. Darusman, 2012. TNF-α Production Inhibitory Activity, Phenolic, Flavonoid and Tannin Contents of Selected Indonesian Medicinal Plants. Research Journal of Medicinal Plants, 6: 406-415.
URL: https://scialert.net/abstract/?doi=rjmp.2012.406.415
URL: https://scialert.net/abstract/?doi=rjmp.2012.406.415
INTRODUCTION
Inflammation is the first response of the immune system to infection or irritation. It is a necessary reaction as a defense mechanism of the human body to eliminate foreign matter (Aguilar-Quesada et al., 2007). It is caused by cytokines such as the tumor necrosis factor-α (TNF-α). The proinflammatory cytokines like reported TNF-α plays a major role in tissue damage (Sullivan, 2004) and inflammatory diseases such as ulcerative colitis (Nikfar et al., 2011), rheumatism (Arend and Gabay, 2004), cardiovascular disease (Salari and Abdollahi, 2011) etc. The inhibition of TNF-α production is can be used to control severe cases of inflammatory diseases and it has become almost commonplace within the past few years (Sullivan, 2004).
Indonesia possesses rich floristic wealth and diversified genetic resources of medicinal plants. It has a wide ranging tropical agro-climatic condition, which are conducive for introducing and domesticating new and exotic plant varieties. The well known Indonesian system of medicine, namely, the Jamu use predominantly plant-based raw materials. A number of Indonesian medicinal plants which are already used for anti-inflammation also exist. Studies about Indonesian medicinal plants had been reported, such as tyrosinase inhibitors and antioxidants (Batubara et al., 2010), oral deodorant (Darusman et al., 2012), antibacterial and antifungal (Goun et al., 2003), antimelanogenesis (Arung et al., 2009), inhibitory activity on nitric oxide (NO) production (Choi and Hwang, 2005), CYP3A4 and CYP2D6 inhibitory activities (Subehan et al., 2006) and anti-acne (Batubara et al., 2009). Since acne and inhibitory activity of NO production are related to inflammation, it is interesting to screen Indonesian medicinal plant as inhibitor of TNF-α production.
Phenolic compounds, which are secondary metabolites in plants, possess a wide spectrum of biological activities such as antioxidant, antimutagenic, anticarcinogenic, as well as anti-inflammatory (Arct and Pytkowska, 2008; Soobrattee et al., 2005). Phenolic compounds include the flavonoid and tannin groups. The aims of this research were to screen the TNF-α inhibitory potency and determine the total phenolic, flavonoid and tannin contents of 28 Indonesian plant species.
MATERIALS AND METHODS
Samples: Indonesian medicinal plants from 20 families were collected from Bogor, Semarang and Samarinda, Indonesia from 2008 until 2009. The identification and voucher specimen of all samples were deposited in Wanariset Samboja, the Wood Anatomy Laboratory, Faculty of Forestry, Mulawarman University, East Kalimantan and Biopharmaca Research Center, Bogor Agricultural University and Herbarium Bogoriense, Bogor, Indonesia.
Extraction: The samples were extracted following the method we previously described (Batubara et al., 2010). Briefly, all samples were dried and grounded before being submitted to methanol (1 g sample: 10 mL solvent) for 12 h for three times. The extract was filtered using Whatman filter paper (No. 1) and concentrated in vacuo at 30°C using a rotary evaporator.
Bioactivity test: In order to determine TNF-α production in response to samples, LPS stimulated cells. Human monocyte-derived THP-1 (ATCC No. TIB 202) cells at the concentration of 1x105 cells mL-1 were cultured in RPMI 1640 medium containing 10% FBS in 24-well plates (450 μL/well) for 3 h at 37°C, 5% CO2. Then the cells were co-incubated with LPS (Escherichia coli 0127 by phenol extraction; Wako 124-05151, less LPS) at 100 ng mL-1 and 50 μL sample at the final concentrations of 1.00 and 10.00 μg mL-1. The cells, LPS and samples were cultured for the next 8 h at 37°C, 5% CO2. The suspension was analyzed for the cytotoxicity. Having been centrifuged, the supernatants were then analyzed for TNF-α production. TNF-α production was determined using a commercial ELISA kit (BIOSOURCE, KHC3011). The cytotoxicity test were determined using WST-1 assay CELL PROLIFERATION ASSAY KIT (Millipore Corporation_C Cat No. 2210, LOT_FJC1619454).
Total phenolic assay: Total polyphenol content was determined using the Folin-Ciocalteu method (Zongo et al., 2010), adapted to microscale. Total polyphenol concentration was calculated from a calibration curve, using gallic acid as the standard (50-800 μg mL-1) and expressed as μg mg-1 gallic acid equivalent using the formula.
Total flavonoid assay: The total flavonoid content was determined as described by Zongo et al. (2010) based on the total content of quercetin, adapted to a microscale. The percentages of flavonoids were calculated from the standard curve of quercetin prepared in methanol and expressed as μg mg-1 quercetin equivalent.
Total tannin assay: Tannin content of samples was determined by a protein precipitation method using Bovine Serum Albumin (BSA) (Wako Pure Chemical Industries, Ltd., Japan). This method was adapted from Kawamoto et al. (1997) by analyzing the BSA content of the supernatant liquid rather than the BSA content of precipitate. Samples were diluted in 50% (v/v) ethanol/water to prepare a concentration of 2 mg mL-1. A 200 μL volume of sample solution was added to 200 μL of BSA solution (10 mg mL-1, dissolved with 0.1 M acetate buffer, pH 5). After reaction at room temperature for 1 h, the solution was centrifuged at 13000 g for 2 min. The remaining BSA in the supernatant was determined by HPLC with a reversed phase Develosil300 C4-HG-5 column (4.6 i.d.x150 mm, Nomura Chemical Co, Ltd, Japan) monitored at 280 nm. The solvent system used was as follow; a linear gradient elution for 20 min from 80 to 20% solvent A (0.01% TFA in water) in solvent B (90% (v/v) CH3CN/water containing 0.01% TFA) at flow rate 1 mL/min. The column temperature was 35°C.
Statistical analysis: All analyses were conducted using the statistical package for the social sciences (SPSS v17.0, Chicago, IL). Descriptive statistics was used to characterize and compare groups. The groups were compared using t-tests and ANOVA for quantitative variables. Significance result was indicated by p value less than five percent (p<0.05).
RESULTS
Extraction: The plant species studied gave methanol extracts with yields ranging from 1.9 to 28.2% based on dried weights of the materials. The highest yield of extract was found on Phaleria papuana leaf (sample No. 18) while the lowest yield was found on Koompassia malaccensis (sample No. 13) (Table 1).
Bioactivity test: Among the 40 plant materials from 28 plant species, 5 plant extracts were toxic to the cell (cell viability lower than 70% at 10 ppm concentration) namely, Helminthostachys zeylanica stem, Intsia palembanica stem, Castanopsis javanica stem, Psidium guajava leaf, Rhizopora sp. stem and Swietenia sp. fruit. About 8 extracts had good cell viability (higher than 90% at 10 ppm concentration) namely Andrographis paniculata herb, Avicennia sp. stem, Goniothalamus macrophyllus leaf, H. zeylanica leaf, Morinda citrifolia leaf, Talinum sp. leaf, Terminalia catappa bark and Vitex pubescens stem. Among 8 plant extracts which had good cell viability, only Avicennia sp. stem and G. macrophyllus leaf methanol extract had high TNF-α inhibitory activity (35.0 and 49.3%, respectively at 10 ppm concentration). The other extracts which had high inhibitory activity were Curcuma domestica rhizome (46.1% at 10 ppm), G. macrophyllus stem (49.3% at 10 ppm) and Guazuma ulmifolia leaf (35.4% at 10 ppm), but these extracts showed lower number of cell viability. Selected data are shown in Fig. 1.
Phytochemical content: The results for total phenolic, flavonoid and tannin contents of the 40 plant materials are presented in Table 1. Total phenolic content ranged from 23.2±1.8 to 1135.0±61.2 μg gallic acid equivalent/mg dry extract.
| Table 1: | Yield, phenolic, flavonoid and tannin contents of all extracts |
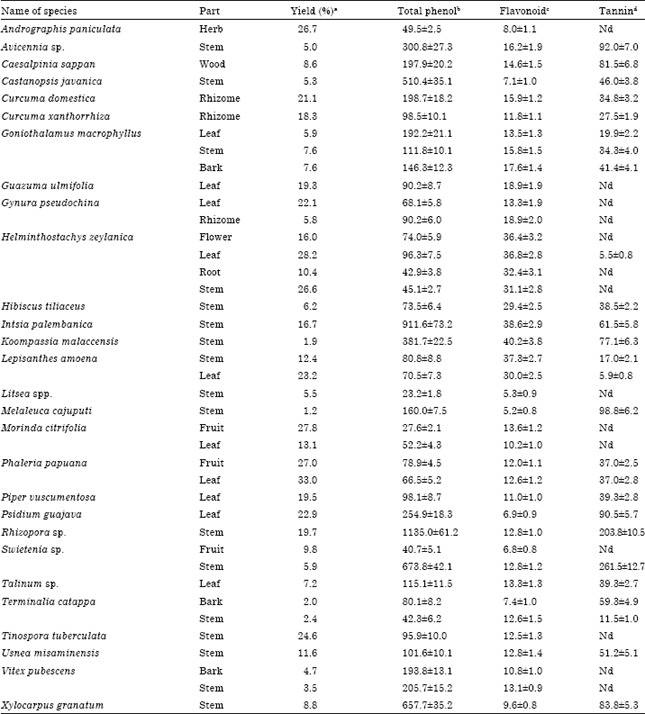 | |
| a: Yield based on dry weight of materials, b: Phenol content is in μg gallic acid equivalent per mg dry extract, c: Flavonoid content is in μg quercetin equivalent per mg dry extract, d: Tannin content is in μg BSA that precipitated per mg dry extract, Nd: Not detected | |
The highest total phenolic content was found in Rhizopora sp. stem followed by I. palembanica stem, Swietenia sp. stem (673.8±42.1 μg gallic acid equivalent/mg dry extract) and Castanopsis javanica stem (510.4±35.1 μg gallic acid equivalent/mg dry extract), while the lowest phenolic content was found in Litsea spp. stem.
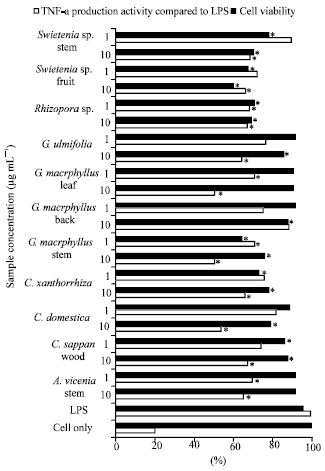 | |
| Fig. 1: | TNF-α production activity and cell viability of selected methanol extracts on THP-1 cells induced by LPS. *: Indicate significant differences p<0.05 |
Total flavonoid content ranged from 5.2±0.8 to 40.2±3.8 μg quercetin equivalent mg-1 dry extract. The highest total flavonoid was found in Koompassia malaccensis (40.2±3.8 μg mg-1) followed by Intsia palembanica (38.6±2.9 μg mg-1). The lowest flavonoid content was found in Melaleuca cajuputi stem (5.2±0.8 μg mg-1) and Litsea spp. stem (5.3±0.9 μg mg-1 which was not significantly different with M. cajuputi stem).
The tannin content of the extracts ranged from undetectable until 261.5±12.7 μg BSA that precipitated mg-1 dry extract. Total tannin was not detected on Andrographis paniculata herb, Guazuma ulmifolia leaf, Gynura pseudochina leaf and rhizome, Helminthotachys zeylanica flower, root and stem, Litsea spp. stem, Morinda citrifolia fruit and leaf, Swietenia sp. fruit, Tinospora tuberculata stem and Vitex pubescens bark and stem. The highest tannin content was found in Swietenia sp. stem (261.5±12.7 μg mg-1) followed by Rhizopora sp. stem (203.8±10.5 μg mg-1).
Correlation between TNF-α production inhibitory activity and phenolic, flavonoid and tannins content: We found no correlation between the TNF-α production inhibitory activity and phenolic, flavonoid and tannins contents of the samples. The highest correlation found was between the TNF-α production inhibitory activity and tannin content with the R2 value of only about 0.1832. The correlation of TNF-α production inhibitory activity and phenolic content was only about 0.0101 (R2), while with flavonoid content was only about 0.0001. The correlation data are shown in Table 2.
| Table 2: | Correlation between TNF-α production inhibitory activity and phenolic, flavonoid and tannin contents |
 | |
DISCUSSION
Extraction: In this study, methanolic extracts of different parts (roots, rhizomes, stems, leaves and fruits) of 28 plants species from different families were screened for their TNF-α production inhibitory activity. The plant families tested in this study were Fagaceae, Myrtaceae, Thymelaeaceae, Acanthaceae, Asteraceae, Menispermaceae, Malvaceae, Piperaceae, Leguminosae, Zingiberaceae, Annonaceae, Sapindaceae, Ophioglossaceae, Rubiaceae, Meliaceae, Portulaceae, Combretaceae, Usneaceae and Verbenaceae.
The extraction was performed by using methanol as the solvent because most of the active compounds were found in methanol extract (Subehan et al., 2006; Batubara et al., 2009). The extract yield of plant materials were varied depending on the plant material used (Table 1, ranging from 1.9 to 28.2% based on dry weight of materials).
Bioactivity test: Many reports had been published for anti-inflammatory active of medicinal plants for example, Tandon and Gupta (2006) reported the effect of Vitex negundo Linn., Gobianand et al. (2010) reported the activity of Cassia fistula Linn. and Meera et al. (2008) reported the activity of polyherbal formulation. These reported data reported were based on in vivo analysis. Another method to analyze the anti-inflammatory effect is by using cells. One report on anti-inflammatory effect used RAW264.7 cells was by Choi and Hwang (2005). They analyzed the NO production inhibitory activity of 75 Indonesian medicinal plants in RAW264.7 cells. Among the 75 plant materials they studied, only 4 plant materials were similar with our study, namely Andrographis paniculata, C. sappan, C. domestica and G. ulmifolia.
Our study was performed in human monocyte-derived THP-1 cells which were induced by lipopolysaccharide (LPS). LPS, a principal component of the outer membrane of gram-negative bacteria is reported to stimulate the production of inflammatory mediator such as TNF-α (Sabina and Rasool, 2007). The bioactivity results of the different parts (roots, rhizomes, stem, leaves and fruits) of 28 Indonesian plant methanol extracts showed that 5 extracts had toxicity against the cells (Fig. 1). The toxicities of these 5 extracts were actually not high. The lowest cell viability was found at 10 ppm of I. palembanica stem extract which only had 51.9% cell viability.
The highest TNF-α production inhibitory activity was found in G. macrophyllus leaf and stem (49.3% at 10 ppm concentration), followed by C. domestica rhizome (46.1% at 10 ppm), G. ulmifolia leaf (35.4% at 10 ppm) and Avicennia sp. stem (35.0% at 10 ppm) (Fig. 1). These results are in agreement with the report of Choi and Hwang (2005) that C. domestica inhibited 66.1% NO production (at 50 ppm concentration), G. ulmifolia leaf inhibited 51.4% NO production (at 50 ppm), A. paniculata did not inhibit NO production at 50 ppm and C. sappan stem only inhibited NO production of about 43.1% at 50 ppm.
Many biological activities of C. domestica rhizome had been reported including anti-inflammatory. This rhizome was reported to have a hepatoprotective effect against paracetamol-induced liver damages in rats (Somchit et al., 2005). The anti-inflammatory report about this species was reported by Rao et al. (1995). On the other hand curcumin as the active component from Curcuma species was also reported to have a protective activity against oxidative stress (Ames et al., 1993).
Focusing on the extract which has good cell viability (not significantly different with control, p<0.05), the results showed that there were only 8 extracts showing good cell viability. Among the 8 extracts, G. macrophyllus leaf methanol extract had the highest TNF-α production inhibitory activity (49.3% at 10 ppm and 28.8% at 1 ppm). The second candidate was Avicennia sp. which had TNF-α production inhibitory activity of about 34.6 at 10 ppm and 30.1 at 1 ppm.
Goniothalamus macrophyllus leaves and stem had higher inhibitory effect on TNF-α production. This species belongs to Annonaceae family and grows in Asia. The biological activities of Goniothalamus species and their metabolites have been described (Blazquez et al., 1999). Interestingly, results showed that G. macrophyllus is one of the sources of drugs for the treatment of cancer and bacterial infection (Wiart, 2007). Present results on G. macrophyllus leaves and stem as TNF-α production inhibitor adds a new bio-activity for G. macrophyllus.
Phytochemical content: The chemical analysis content was performed to find the prospective chemical group which is responsible for the biological activity. Total phenolic and flavonoid contents of extracts were determined spectrophotometrically, while tannin content was determined by High Performance Liquid Chromatography (HPLC). The phenolic, flavonoid and tannin contents of all extracts are shown in Table 1. Phenolic , flavonoid and tannin previously reported had relationship with inhibitory activity especially on the enzyme inhibitory activity (Ji and Zhang, 2006).
The amount of phenolic, flavonoid and tannin compounds varied from different plant species. The composition of the extracts and the yields of phenol, flavonoid and tannin were found to be dependent on the plant species, the plant part utilized and the type of plant material (woody or herb). The plants studied gave methanol extracts with yields ranging from 1.92 to 28.21%. The highest phenolic content was found in the woody plants like Rhizopora sp., I. palembanica, Swietenia sp. and C. javanica. The herbal plants showed lower phenolic content compared to woody plants.
Flavonoids, one group of phenolic compounds, were found in all extracts. Plant flavonoids have local action on some diseases like inflammation, oedema, pain, fungal infections etc (Amit et al., 2007). Flavonoid contents ranged from 5.2 to 40.2 μg quercetin equivalent/ mg dry extract. The highest flavonoid content was also found in woody plants with high phenolic content. Like flavonoid content, tannin content was also found high in woody plants compared to herbal plants. Tannin content could not be detected in some herbal plants.
The phenolic, flavonoid and tannin contents had no correlation with TNF-α production inhibitory activity of extracts. A very low negative correlation was found between tannin content and TNF-α production inhibitory activity (R2 = 0.1832). It made suggestion that the active component would not be tannin compound. In Goniothalamus species, some compounds have been isolated such as naringenin, goniothalamin and pinocembrine (Fig. 2) (Ahmad et al., 2010). Between the three compounds, naringenin had been reported to have an effect as TNF-α inhibitor (Hirai et al., 2007). Our next study will deal with the isolation of active components from Goniothalamus macrophyllus leaf which could have TNF-α production inhibitor activity.
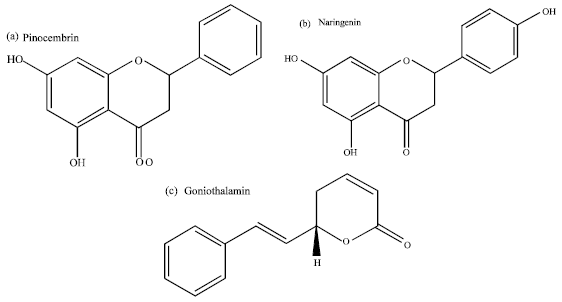 | |
| Fig. 2: | Structure of compounds in Goniothalamus species |
CONCLUSION
Among the 40 Indonesian medicinal plant materials, Goniothalamus macrophyllus leaf methanol extract was the most active extract as TNF-α production inhibitor with high cell viability. The phenolic, flavonoid and tannin contents of samples had no correlation with TNF-α production inhibitory activity.
ACKNOWLEDGMENT
We gratefully acknowledge the preparation of samples by the Biosfer Manusia foundation, East Kalimantan; Biopharmaca Research Center, Bogor Agricultural University, Bogor, West Java and Faculty of Forestry Mulawarman University, Samarinda, East Kalimantan, Indonesia. We also would like to thank the Directorate General of Higher Education, Republic of Indonesia for Program of Academic Recharging-C to IB in manuscript preparation 2011.
REFERENCES
- Aguilar-Quesada, R., J.A. Munoz-Gamez, D. Martin-Olivia, A. Peralta and M.T. Valenzuela et al., 2007. Interaction between ATM and PARP-1 in response to DNA damage and sensitization of ATM deficient cells through PARP inhibition. BMC Mol. Biol., 8: 29-29.
CrossRef - Ames, B.N., M.K. Shigenaga and T.M. Hagen, 1993. Oxidants, antioxidants and degenerative diseases of aging. Proc. Natl. Acad. Sci., 90: 7915-7922.
CrossRef - Amit, G., M.S. Ashawat, S. Shailendra and S. Swarnlata, 2007. Phytosome: A novel approach towards functional cosmetics. J. Plant Sci., 2: 644-649.
CrossRefDirect Link - Arct, J. and K. Pytkowska, 2008. Flavonoids as components of biologically active cosmeceuticals. Clin. Dermatol., 26: 347-357.
CrossRefPubMedDirect Link - Arend, W. and C. Gabay, 2004. Cytokines in the rheumatic diseases. Rev. Rheum. Dis. Clin. North Am., 30: 41-67.
PubMed - Arung, E.T., I.W. Kusuma, E.O. Christy, K. Shimizu and R. Kondo, 2009. Evaluation of medicinal plants from Central Kalimantan for antimelanogenesis. J. Nat. Med., 63: 473-480.
CrossRefDirect Link - Batubara, I., T. Mitsunaga and H. Ohashi, 2009. Screening anti-acne potency of Indonesian medicinal plants: Antibacterial, lipase inhibition and antioxidant activities. J. Wood Sci., 55: 230-235.
CrossRefDirect Link - Batubara, I., L.K. Darusman, T. Mitsunaga, M. Rahminiwati and E. Djauhari, 2010. Potency of Indonesian medicinal plants as tyrosinase inhibitor and antioxidant agent. J. Biol. Sci., 10: 138-144.
CrossRef - Blazquez, M.A., A. Bermejo, M.C. Zafra-Polo and D. Cortes, 1999. Styryl-lactones from Goniothalamus species-A review. Phytochem. Anal., 10: 161-170.
CrossRef3.0.CO;2-2/abstract target='_blank' class='btn btn-sm btn-outline-primary mr-3 mt-3'>Direct Link - Choi, E.M. and J.K. Hwang, 2005. Screening of Indonesian medicinal plants for inhibitor activity on nitric oxide production of RAW264.7 cells and antioxidant activity. Fitoterapia, 76: 194-203.
CrossRefPubMedDirect Link - Darusman, H.S., M. Rahminiwati, S. Sadiah, I. Batubara, L.K. Darusman and T. Mitsunaga, 2012. Indonesian Kepel fruit (Stelechocarpus burahol) as oral deodorant. Res. J. Med. Plants, 6: 180-188.
CrossRefDirect Link - Gobianand, K., P. Vivekanandan, K. Pradeep, C.V.R. Mohan and S. Karthikeyan, 2010. Anti-inflammatory and antipyretic activities of indian medicinal plant Cassia fistula Linn. (golden shower) in wistar albino rats. Int. J. Pharmacol., 6: 719-725.
CrossRefDirect Link - Goun, E., G. Cunningham, D. Chu, C. Nguyen and D. Miles, 2003. Antibacterial and antifungal activity of Indonesian ethnomedical plants. Fitoterapia, 74: 592-596.
CrossRefDirect Link - Hirai, S., Y. Kim, T. Goto, M.S. Kang, M. Yoshimura, A. Obata, R. Yu and T. Kawada, 2007. Inhibitory effect of naringenin chalcone on inflammatory changes in the interaction between adipocytes and macrophages. Life Sci., 81: 1272-1279.
CrossRef - Ji, H.F. and H.Y. Zhang, 2006. Theoretical evaluation of flavonoids as multipotent agents to combat Alzheimer's disease. J. Mol. Struct. THEOCHEM, 767: 3-9.
CrossRef - Kawamoto, H., K. Mizutani and F. Nakatsubo, 1997. Binding nature and denaturation of protein during interaction with galloylglucose. Phytochemistry, 46: 473-478.
CrossRef - Meera, S., N.S. Kumar and V.S.S.S. Guptatyam, 2008. Screening of anti-arthritic, anti-inflammatory and analgesic activity of a polyherbal formulation. Int. J. Pharmacol., 4: 398-402.
CrossRefDirect Link - Nikfar, S., S. Ehteshami-Afshar and M. Abdollahi, 2011. A systematic review and meta-analysis of the efficacy and adverse events of infliximab in comparison to corticosteroids and placebo in active ulcerative colitis. Int. J. Pharmacol., 7: 325-332.
CrossRef - Salari, P. and M. Abdollahi, 2011. A comprehensive review of the shared roles of inflammatory cytokines in osteoporosis and cardiovascular diseases as two common old people problem; Actions toward development of new drugs. Int. J. Pharmacol., 7: 552-567.
CrossRefDirect Link - Sabina, E.P. and M. Rasool, 2007. Therapeutic efficacy of indian ayurvedic herbal formulation triphala on lipid peroxidation, antioxidant status and inflammatory mediator TNF- α in adjuvant-induced arthritic mice. Int. J. Biol. Chem., 1: 149-155.
CrossRefDirect Link - Soobrattee, M.A., V.S. Neergheen, A. Luximon-Ramma, O.I. Aruoma and T. Bahorun, 2005. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res./Fundam. Mol. Mech. Mutagen., 579: 200-213.
CrossRefDirect Link - Somchit, M.N., A. Zuraini, A.A. Bustamam, N. Somchit, M.R. Sulaiman and R. Noratunlina, 2005. Protective activity of turmeric (Curcuma longa) in paracetamol-induced hepatotoxicity in rats. Int. J. Pharmacol., 1: 252-256.
CrossRefDirect Link - Subehan, T., Usia, H. Iwata, S. Kadota and Y. Tezuka, 2006. Mechanism-based inhibition of CYP3A4 and CYP2D6 by Indonesian medicinal plants. J. Ethnopharmacol., 105: 449-455.
Direct Link - Sullivan, K.E., 2004. TNF and TNF-αinhibitors: Mechanism of action. Pediatric Rheumatol. Online J., 2: 7-22.
Direct Link - Tandon, V.R. and R.K. Gupta, 2006. Anti-inflammatory activity and mechanism of action of Vitex negundo Linn. Int. J. Pharmacol., 2: 303-308.
CrossRefDirect Link - Zongo, C., A. Savadogo, L. Ouattara, I.H.N. Bassole and C.A.T. Ouattara et al., 2010. Polyphenols content, antioxidant and antimicrobial activities of Ampelocissus grantii (Baker) Planch. (Vitaceae): A medicinal plant from Burkina Faso. Int. J. Pharmacol., 6: 880-887.
CrossRefDirect Link - Wiart, C., 2007. Goniothalamus species: A source of drugs for the treatment of cancers and bacterial infections? Evidence-Based Complementary Altern. Med., 4: 299-311.
CrossRefDirect Link