
Fatih Seyis
Not Available
Wolfgang Friedt
Not Available
Axel Voss
Not Available
Wilfried Luhs
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 5 | Page No.: 593-596







ABSTRACT
In the course of studies dealing with the inheritance of erucic acid content in the seed oil of Brassica species individual plants belonging to two accessions of B. oleracea conv. capitata, Kashirka 202 and Ladozhskaya DS 8395, were identified being very low in erucic acid content. Applying half seed selection their progeny confirm a monogenous inheritance. Low-erucic mutants of B. oleracea have been found and described first time.
PDF Abstract XML References Citation
How to cite this article
Fatih Seyis, Wolfgang Friedt, Axel Voss and Wilfried Luhs, 2004. Identification of Individual Brassica oleracea Plants with Low Erucic
Acid Content. Asian Journal of Plant Sciences, 3: 593-596.
DOI: 10.3923/ajps.2004.593.596
URL: https://scialert.net/abstract/?doi=ajps.2004.593.596
DOI: 10.3923/ajps.2004.593.596
URL: https://scialert.net/abstract/?doi=ajps.2004.593.596
INTRODUCTION
Due to substantial progress in breeding and cultivation practice rapeseed and mustards-derived from several locally distributed members of the genus Brassica-have become one of the worldwide most important source of vegetable oil. Especially in several European countries with cool-temperate climates oilseed rape (B. napus) with ‘double-low‘ seed quality (canola) dominates field crop production.
Regarding this quality characters, low-erucic acid mutants were found in B. rapa (AA)[1], B. napus (AACC)[2,3] and B. juncea (AABB)[4].
In the amphidiploid B. carinata (BBCC) low-erucic acid mutants were not known up to the 90′s. Different strategies, such as the selection of transgressive segregants in the crossing progeny of diverse B. carinata accessions[5], induced mutations and interspecific crosses with low-erucic B. napus and B. juncea[6,7] were used to reduce the erucic acid content. With the first and last mentioned method low erucic acid forms could be selected successfully[5,8].
For the monogenomic species B. oleracea and B. nigra low-erucic acid forms were not cited in the literature up to now[1-3]. In the course of studies dealing with the inheritance of erucic acid content in the seed oil of Brassica species individual plants belonging to two accessions of B. oleracea conv. capitata, Kashirka 202 and Ladozhskaya DS 8395, were identified being very low in erucic acid content. In the presented data their morphological characters and their fatty acid composition were described.
MATERIALS AND METHODS
Plant material: Individual plants belonging to two accessions of B. oleracea conv. capitata, Kashirka 202 and Ladozhskaya DS 8395, were identified being very low in erucic acid content in 1999 in the Plant Breeding Institute of the Justus-Liebig-University of Giessen. Individual half seed plants were transferred in pots and plant characters like leaf characteristics, flower characteristics were obtained. They were cultivated in the greenhouse, artificially vernalised and self-pollinated by bagging. The seed progeny was analysed for fatty acid composition in order to check for true-breeding individuals.
The fatty acid composition of the B. oleracea material was determined on the basis of half-seed analysis as described earlier using GC[9,10]. The preparation of half seeds was done according to the method described by Thies[11].
RESULTS
Description of investigated B. oleracea forms: Kashirka and Ladozhskaya both have vernalization requirement. After vernalization experiments it can be said that Kashirka has a vernalization requirement of 14 weeks at 5°C and Ladozshkaya up to 18 weeks at 5°C.
Both genotypes are self-compatible and seed set could be obtained after pollination per hand (Fig. 1a and b). Their flowers are typical Brassica flowers (Fig. 1c).
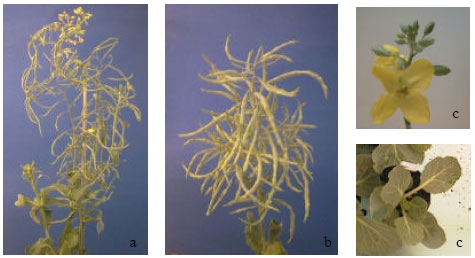 | |
| Fig. 1(a-d): | Seed sed of Ladozhskaya (a1) and Kashirka (1b), typical Brassica flower (1c) and leaf shape (1d) |
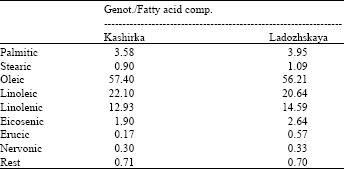
| Table 1: | Mean fatty acid composition (%) of individual half seed plants belonging to the two genotypes Kashirka and Ladozhskaya (mean of totally 21 and 23 plants, respectively) |
 | |
One stigma, 4 long and two short anthers, 4 sepals and 4 petals. The leaves of both accessions are small and transverse broad elliptic (Fig. 1d).
Quality characters
Fatty acid composition: After half seed analysis 23 individuals of Kashirka and 21 individuals of Ladozshkaya were identified with low erucic acid content.
The mean values of the fatty acid composition of these half seed plants can be seen in Table 1. The fatty acid composition of both cabbages is very similar. The biggest component for Kashirka and Ladozskaya is oleic acid, (respectively 57.40 and 56.21%), followed by linoleic acid (22.10 and 20.64%, respectively) and linolenic acid (12.93 and 14.59%, respectively). Low amounts of palmitic, stearic and erucic acid were also detected. Specially erucic acid is under the 2% limit given for oils regarding canola oil quality.
Unfortunately only three half seed plants of Kashirka and one half seed plant of Ladohszkaya survived and could be selfed and they also display low erucic acid content. The obtained seed material was measured with NIRS (Near Infrared Restriction Spectroscopy) for glycosinolate content. These values are 63.4 and 129.5 μmol g/DM, respectively. Regarding Canola quality they have only O-quality.
Addittionally to the presented results the genomic Southern analysis and PCR-Amplifications of different Brassica species inclusively Ladozhskaya revealed that in the progeny of the B. oleracea accession Ladozshkaya DS 8395 a complete different pattern was observed as no signal in the selection zero of this cabbage meaning partial or complete deletion of the KCS gene. The latter result was verified through PCR analysis with a KCS-specific primer pair, where the 1.25 kb fragment characteristics for all other genotypes was not amplified in the case of zero erucic acid mutant[12].
DISCUSSION
The gene pool of elite rapeseed breeding material has been further eroded by an emphasis on specific quality traits. As a consequence, genetic variability in this important oilseed crop is restricted with regard to many characters of value for breeding purposes. Resynthesis of novel genotypes with novel quality characters through artificial crosses between the diploid parents, assisted by embryo rescue techniques, will broaden the genetic basis of present breeding material.
Developing synthetic Brassica napus forms has provided important basic germplasm for further improvements of seed yield, disease and pest resistance as well as relevant seed quality traits[13]. Regarding the later some examples of introgression breeding are documented, such as the resynthesis approach to create new genetic variation for low glucosinolate content[14,15] or the approaches developing of yellow-seeded rapeseed via interspecific crosses[16,17]. Regarding fatty acid composition different cauliflower cultivars (B. oleracea ssp. oleracea convar. botrytis var. botrytis) and Yellow Sarson (B. rapa ssp. trilocularis) were used in an earlier study for wide hybridisation to create novel rapeseed with high erucic acid content (HEAR) and which is now used as basic material for the improvement of industrial rapeseed[12,18-20]. Similar experiments were conducted by Chen and Heneen[13], Health and Earle21,22], Rahman et al.[23], Lu et al.[24] and Rahman[25] using interspecific Brassica hybridization in order to modify the fatty acid composition of oilseed rape.
In fact, current double-low breeding material seems to be closely, related and intensive quality breeding-using the spring rapeseed cultivars Liho/Oro (low erucic acid source) and the Polish cultivar Bronowski (low glycosinolate source) as unique donors for quality improvement-has also contributed to narrow the genetic base of oilseed rape[26,27].
The limit for low glycosinolate content in B. napus is 30 μmol/100 g DM in rapeseed cultivars of the EU[28]. The glucosinolate content of the low-erucic-acid mutants are about this limit. But if we use these mutants in interspecific crosses with very low gylcosinolate B. rapa forms, the content of the obtained hybrids will be probably under this limit.
Therefore, the discovery of low-erucic acid mutants among the mentioned B. oleracea accessions and the development of synthetic rapeseed forms via interspecific crosses with interesting 0-or 00-quality B. rapa genotypes will open further the possibility to use RS-rapeseed material as a genetic resource for example for quality and yield improvement in Brassica napus, perhaps in developing RS-lines with 0-or 00-quality.
REFERENCES
- Fernandez-Escobar, J., J. Dominguez, A. Martin and J.M. Fernadez-Martinez, 1988. Genetics of the erucic acid content in interspecific hybids of Ethiopian Mustard Brassica carinata Braun) and rapeseed B. napus L.). Plant Breed., 100: 310-315.
Direct Link - Getinet, A., G. Rakow, J.P. Raney and R.K. Downey, 1994. Development of zero erucic acid Ethiopian mustard through an interspecific cross with zero erucic acid Oriental mustard. Can. J. Plant Sci., 74: 793-795.
Direct Link - Chen, B.Y. and W.K. Heneen, 1992. Inheritance of seed colour in Brassica campestris L. and breeding for yellow seeded B. napus L. Euphytica, 59: 157-163.
CrossRefDirect Link