
Neveen S.I.y Geweely
Cairo University, Giza, Egypt
Biotechnology
Year: 2006 | Volume: 5 | Issue: 3 | Page No.: 358-364







ABSTRACT
Extraction and purification of the intracellular urease of Rhizopus oryzae was carried out through different steps. Production and crushing the collected cells and preparation of the cell free homogenate were estimated. The intracellular R. oryzae urease was purified to homogeneity by salting out with ammonium sulphate, dialysis and passage through chromatography resins (Sephdex G-200 column, Sephdex G-100 and Diethylaminoethyl cellulose column) and test for purity by simple polyacrylamide gel electrophoresis technique. The purified enzyme resulted in 622 fold by native molecular size of 172 kDa, exhibited a specific activity in the presence of urea of 112 U mg-1 protein with the recovery of 26%. Studying factors affecting the activity of the purified urease enzyme resulted in: There was a proportional increase of enzyme activity corresponding to the increase of the enzyme concentration. The optimum concentration of urea was (20 μg mL-1). An optimum temperature was 55oC. The purified enzyme was most active at pH 7. Potassium, zinc and cupper salts, each inhibited activity of the purified enzyme. Urease activity was enhanced by the presence of magnesium, manganese and calcium salts, but inhibited completely by mercury. The reversible urease inhibitors, ferrous and cobalt salts, blocked enzyme activity in the crude mycelial fraction when added at a concentration of 10-3 M.
PDF Abstract XML References
How to cite this article
Neveen S.I.y Geweely, 2006. Purification and Characterization of Intracellular Urease Enzyme Isolated from Rhizopus oryzae. Biotechnology, 5: 358-364.
DOI: 10.3923/biotech.2006.358.364
URL: https://scialert.net/abstract/?doi=biotech.2006.358.364
DOI: 10.3923/biotech.2006.358.364
URL: https://scialert.net/abstract/?doi=biotech.2006.358.364
INTRODUCTION
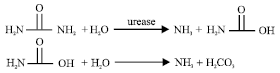
Urease (urea amidohydrolase, EC 3.5.1.5) is a nickel-containing enzyme that catalyzes the hydrolysis of urea to yield ammonia and carbamat, the latter compound decomposes spontaneously to generate a second molecule of ammonia and carbon dioxide (Andrews et al., 1986; Hausinger, 1993; Park and Hausinger, 1995; Pearson et al., 1997). Urease has many industrial applications in diagnostic kits for measuring urea, in alcoholic beverages as a urea reducing agent (Fujinawa and Dela, 1990; Fumuyiwa and Ouch, 1991). Urea's commercial uses for the manufacture of plastics, hair conditioners, glues, fertilizer and animal feed as a browning agent in factory-produced pretzels and also drug uses of urea in topical dermatological products, for nonsurgical debridement of nails and as a diuretic (Wikipedia, 2006). There are many microbial sources for this enzyme including bacteria such as Lactobacillus ruminis, Lactobacillus fermentum and Lactobacillus reuteri (Kakimoto et al., 1989; Kakimoto and Suzuki, 1992) and fungi such as Rhizopus oryzae (Farley and Santosa, 2002). Filamentous fungi are the sources of about 40% of all available urease enzymes (Archer and Peberdy, 1997). It is also the industry preference to use filamentous fungi, as enzyme producers, because the fermentation industries are very familiar with the conditions required to maximize production of the homologous proteins by them (Wubbolts et al., 2000). Molecular biology of microbial urease was studied by Mobely et al. (1995). Purification and Properties of the Klebsiella aerogenes urease was studied by Mulrooney et al. (2005). Microbial urease was studied by Remaut et al. (2001), Mehta et al. (2003), Won and Lee (2004) and Zambelli et al. (2005).
 |
The purpose of present study was to study intracellular urease production and purification from Rhizopus oryzae and some physicochemical parameters were tested and to find a purified urease for possible use in diagnostic kits of urea measurement in medical laboratories.
MATERIALS AND METHODS
Organism used: Rhizopus oryzae a local isolate from an Egyptian soil was studied for urease production.
Culture conditions: The medium contained: urea 2.26 g, glucose 20 g, MgSO4.7H2O 0.5 g, potato infusion 4 g, bacto phenol-red 0.12 g and distilled water up to 1 L. The medium was dispensed to 250 mL Erlenmeyer flasks at the rate of 50 mL per flask. Each flask was inoculated with a fungal disc (1 cm diameter) taken from the colony margin of 2-4 day old cultures. Triplicate flasks were incubated at 27 oC for 10 days Smith et al. (1993).
Preparation of intracellular urease crude enzyme (cell free extract): Mats from liquid cultures were harvested by centrifugation at 3,000 rpm for 5 min and then washed with 0.1 M potassium phosphate buffer (pH 8.0) and vortexes twice together with an equal volume of glass beads (0.5 mm diameter ) for 5 min at 4oC. The disintegrated cell mixture was centrifuged at 13,000 rpm for 5 min at 4oC (Walfridsson et al., 1997). The supernatant was stored in refrigerator until analyzed for urease enzyme activities.
Enzyme assay: In the semi-quantitative screening method, the enzyme production was evaluated by visual inspection of the phenol red color change, when enzyme exists, the color changes from red to pink caused by ammonia liberation in the culture tubes. The amount of color change intensity was correlated to the enzyme production. In the quantitative urease assay, the enzyme activity in media supernatants was measured by Weatherburn (1967) with some modifications, i.e., Na2 HPO4 was used instead of NaOH in alkaline hypochlorite solution and the 20 min time for color development was elongated to 30 min. The reactions were done in micro tubes containing 100 μL of sample, 500 μL of 50 mM urea and 500 μL of 100 mM potassium phosphate buffer (pH 8.0) in a total volume of 1.1 mL. The reaction mixture was incubated in a shaking water bath at 37°C for 30 min. The reaction stopped by transferring 50 μL of reaction mixture to the tubes containing 500 μL of phenol-sodium nitroprusside solution (0.05 g sodium nitroprosside + 1 g phenol/100 mL distilled water). Five hundred micro liters of alkaline hypochlorite (3.56 g Na2HPO4 + 1 mL sodium hypochlorite + 100 mL distilled water) was added to the tubes and incubated at room temperature for 30 min. Finally, the optical density of the color complex was measured at 630 nm and compared to a standard curve prepared with (NH4)2SO4. One unit of urease activity was defined as the amount of enzyme liberating 1 μmol NH3 from urea per minute, under the standard assay conditions.
Protein determination: Protein concentration was determined spectrophotometrically at 260 nm by the method of Segel (1968).
Purification of the intracellular urease: A complete purification process were carried out as follow: Ammonium sulphate fractionation (at 80% saturation) followed by gel filteration using Sephdex G-200 column (38.5x2.25 cm) and then Sephdex G-100 column (10x1.25 cm), finally preparation of diethylaminoethyl cellulose column (DEAE) (1.6x55 cm) were performed according to Kimikasu et al. (1967).
Estimation of the molecular weight of the purified enzyme: Sodium dodecyl-sulphatepolyacrylamide gel electrophoresis (SDS-PAGE) was carried out on 12.5% acrylamide gel according to the method of Laemmli (1970). Urease was visualized by silver staining as described by Wray et al. (1981). A protein sample (12 μL) containing about 10-15 μg protein was mixed with 3 μL of sample buffer and heated in boiling water bath for 5 min followed by centrifugation. The molecular mass standards (catalase, 232 kDa; aldolase, 158 kDa and bovine serum albumin, 65 kDa) were used.
Factors affecting the activity of the purified urease: The following factors were investigated for their effect on the activity of the purified urease enzyme preparation: different volumes (μg mL-1) of the purified enzyme on its activity and urea at different concentrations, respectively. Different temperatures according to Bongaerts et al. (1978), different pH values and various metal salts according to Kimikasu et al. (1967).
RESULTS AND DISCUSSION
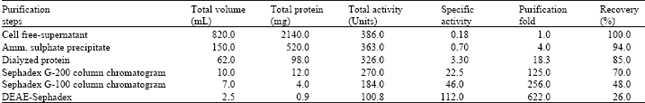
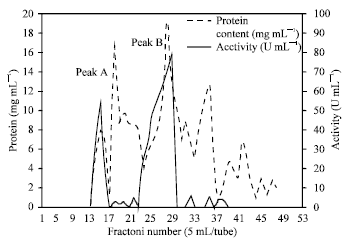
Purification of the intracellular urease: The cell free-supernatant urease enzyme was subjected to a purification procedure consisting of 5 steps. The specific activity of urease in the crude extract was 0.18 U mg-1 proteins Table 1. By using ammonium sulfate precipitation, 4.0 fold increase in specific activity was manifested while the recovered activity was 94.0%. From the elution profile of Sephadex G-200 column chromatography in Fig. 1 and Table 1, it can be seen that R. oryzae intracellular urease was eluted from the column in the two peaks of urease activity accompanied with several peaks of protein. The first peak (A) of activity was found in the fractions (11-15) while the second peak of activity (B) was found in the fractions (21-29).
Only fractions of peak No. (A) with the highest urease specific activity were pooled and were collected separately in the form of one fraction, precipitated again with ammonium sulphate at 80% concentration and then centrifuged at 20,000 rpm for 30 min.
| Table 1: | Purification scheme of Rhizopus oryzae intracellular urease enzyme |
 | |
 | |
| Fig. 1: | Typical elution profile for the behavior of R. oryzae intracellular urease on Sephadex G-200 |
 | |
| Fig. 2: | Typical elution profile for the behavior of R. oryzae intracellular urease on Sephadex G-100 |
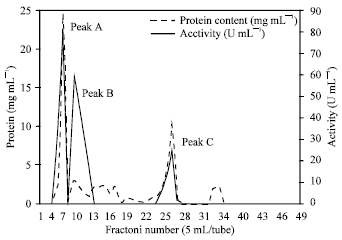
The resulting precipitate was dissolved in a minimum amount of potassium phosphate buffer (pH 8.0), concentrated and fractioned again through a column of Sephadex G-100 previously equilibrated with the same buffer and applied with 12 mg protein of the previously prepared purified urease solution.
The active fraction of peak No. B obtained after the 2nd purification step Fig. 2 were collected and concentrated by dialysis against sugar crystals. Seven mL were obtained and applied (4 mg protein) to DEAE-Sephadex column and the eluate was carried out using 0.05 M potassium phosphate buffer (pH 8.0) with a step-wise gradient from 0.0-0.2 M NaCl prepared in the same buffer.
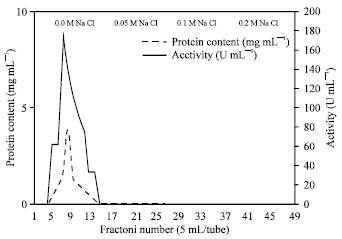 | |
| Fig. 3: | Typical elution profile for the behavior of R. oryzae intracellular urease peak B on DEAE-Sephadex |
Fractions were collected and assays of the enzyme activity and protein concentration were done for all fractions. Finally, the specific activity with the purified intracellular urease enzyme was 112 U mg-1 protein an enrichment of about 622 fold and recovery of 26% Table 1 and Fig. 3. This specific activity is higher than that observed for most cyanobacterial ureases purified (Carvajal et al., 1982; Rai, 1989). Urease production was investigated among thirteen strains of Aspergillus niger isolated from soils in Iran where the maximum enzyme productions were 106 and 109 U gm-1 dry mass (Ghasemi et al., 2004). The first analysis of urease enzyme in Rhizopus oryzae (Zygomycetes ) was investigated by Farley and Santosa (2002), where the regulation of intracellular urease activity (2.4 U mg-1 protein) was examined. Mirbod et al. (2002) and Lubbers et al. (1996) studied urease production by a pathogenic fungus Coccidioides immitis and Schizosaccharomyces pombe with the total activity of 3340 and 1199 U L-1, respectively. The specific activities of urease up to 180000 U mg-1 protein were observed by Mobley and Hausinger (1989). The urease from Prochlorococcus marinus was purified 900 fold to a specific activity of 94.6 μmol urea/min/mg protein by liquid chromatography methods (Palinska et al. 2000).
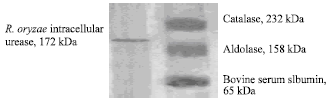 | |
| Fig. 4: | Silver-strained PAGE of the purified R. oryzae intracellular urease |
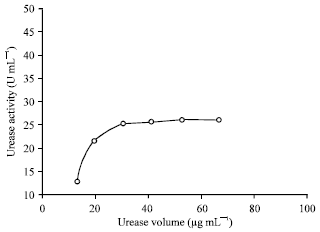 | |
| Fig. 5: | Effect of different volumes (μg mL-1) of the purified R. oryzae intracellular urease on its activity. LSD at 0.05 = 4.8 |
Urease has been purified by DEAE-cellulose column chromatography and the specific activity of the purified enzyme was 1920 U mg-1 with the recovery of 8% (Pandey and Pandey, 1991). Four fungal species, Aspergillus awamori, A. flavus, Penicillium chrysogenum and Trichoderma koningii showed the highest rates of urease enzyme formation (Omar and Abd-Alla, 2000). The purification of urease from Aspergillus nidulans was studied by Mackay and Pateman (1980), Creaser and Porter (1985). The regulation of urease activity in Aspergillus nidulans was found by Mackey and Pateman (1982). Smith et al. (1993) recorded the isolation and characterization of urease from Aspergillus niger. The tested active fractions obtained after the 3rd purification step were collected and used for the next experiment.
Estimation of the molecular weight of the purified urease produced by R. oryzae: The molecular weight of the purified intracellular urease was determined by electrophoresis using sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE). The estimated molecular weight of R. oryzae intracellular urease was 172 kDa Fig. 4. This structure yields a predicted native molecular mass of R. oryzae urease makes it the smaller urease purified so far. The result coupled with that found by Palinska et al. (2000) where the molecular mass was 168 kDa of the Prochlorococcus marinus urease.
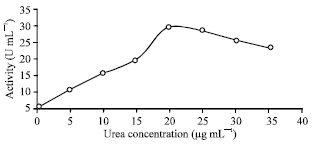 | |
| Fig. 6: | Effect of different concentration of urea (acting substrate) on the activity of the purified R. oryzae intracellular urease. LSD at 0.05 = 6.4 |
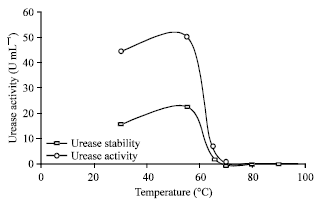 | |
| Fig. 7: | Temperature dependence of the activity and stability of R. oryzae intracellular urease. LSD at 0.05 = 2.41 |
Within the lower range are the molecular masses of urease of Anabaena cylindrica (197 kDa) Argall et al. (1992) and Spirulina maxima (232 kDa) Carvajal et al. (1982). The apparent molecular masses for ureases vary from 200 to 360 kDa, the highest being 800 kDa Mobley and Hausinger (1989). The native molecular size of Morganella morganii urease was 590 kilodaltons Hu et al. (1990). The molecular mass of Staphylococcus leei urease was 480 kDa (Jin et al., 2004).
Two criteria for the purity of the urease enzyme from R. oryzae were testimonized. DEAE-Sephadex column chromatography (final stage of purification) resulted in a single sharp peak (Fig. 3). The second was given by applying SDS-PAGE electrophoresis technique that resulted in one protein spot.
Characterization of pure R. oryzae urease: There was a continuous increase of urease activity by the increases of its volume in the reaction mixture. This was true up to 30 μg mL-1 of enzyme volume Fig. 5 which is in accordance with the general behavior of most enzymes. The optimum urea concentration (acting substrate) in the reaction mixture was 20 ug mL-1 beyond which no obvious change was observed Fig. 6. A lower urease activity was observed in ammonium-grown cells than in the presence of urea or nitrate (Collier et al., 1999).
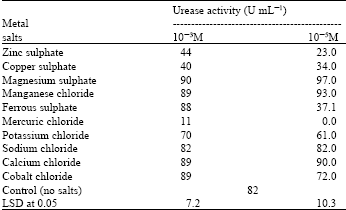
| Table 2: | Effect of metallic salts on the activity of the purified urease enzymes |
 | |
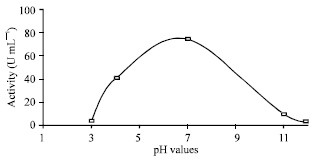 | |
| Fig. 8: | Effect of different pH values on the purified R. oryzae intracellular urease. LSD at 0.05 = 4.1 |
Different utilizable nitrogen sources (ammonium, urea or alanine) exhibit a urease activity (Ge et al., 1990).
The temperature stability was determined after a preincubation of the purified enzyme for 15 min at the temperature indicated and rapid cooling at 30°C. Maximum activity was obtained at 55°C. The stabilization of the enzyme may be favored at high protein concentration, since the protein concentration in the cell-free extract was approximately 2140 mg mL-1, whereas in the assay with the purified enzyme, protein concentration was approximately 0.9 mg mL-1. The enzyme was stable for at least 15 min at temperatures between 40 and 60°C, while above 60°C, a rapid irreversible inactivation occurred Fig. 7. Similarly, Jahns (1995) stated that the highest urease activities of Sporobolomyces roseus on media containing urea was heat-stable and showing no loss of activity after incubation at 70°C for 15 min.
The recorded optimum pH of 7 is very close to value 7.4 which reported for Helicobacter Pylori Urease by Singh (1990) and Jahns et al. (1995). The pure enzyme was stable in the range between pH 4A0 and 11A0 in three different buffer systems (50 mM citrate, 50 mM phosphate and 50 mM diethyl-barbiturate). Below or above these pH values, an irreversible inactivation was observed Fig. 8.
Table 2 indicate that the inhibition of the tested purified urease by potassium, zinc and cupper salts, where as the complete inhibition of the tested purified urease by mercury was in agreement with that obtained by Rai and Singh (1987) and Mobley et al., (1995), which has been suggested to block ammonium uptake (Singh et al. 1983), while dissimilar to that recorded for urease of Prochlorococcus marinus (Palinska et al. 2000). The inhibitor may symmetrically bridging the two Ni ions in the active site through the hydroxamate oxygen and chelating one Ni ion through the carbonyl oxygen and the flexible flap flanking the active site cavity of urease enzyme (Rypniewski et al. 2000). On the other hand, the tested purified urease was activated by all concentrations of magnesium, manganese and calcium salts, which in accordance with Mirbod et al. (2002) who stated that urease enzyme from pathogenic fungus was enhanced by the presence Mg2+ or Mn2+, but inhibited by Li+, Ni2+, Cu2+ or Zn2+. Only stimulating effect on the tested purified urease was showed at the low concentration (10-3) of Ferrous and Cobalt salts but inhibitory at high concentration (10-5). Sodium chloride exerted no effect at all concentrations. Omar and Abd-Alla (2000) revealed that Cd2+, Hg2+ and Zn2+ generally inhibited urease enzyme activity while Cu1+, Fe3+ and Al3+ showed a stimulatory effect. Inhibition of urease by acetohydroxamate was recorded by Rai and Singh (1987). The inhibition of urease by amides and esters of phosphoric acid was detedcted by Andrews et al. (1986). Physiologic Characterization of urease of Actinomyces naeslundii were tested by Bermudez and Burne (1999).
REFERENCES
- Andrews, R.K., A. Dexter, R.L. Blakeley and B. Zener, 1986. Jack bean urease (EC 3.5.1.5). On the inhibition of urease by amides and esters of phosphoric acid. J. Am. Chem. Soc., 108: 7124-7125.
Direct Link - Archer, D.B. and J.F. Peberdy, 1997. The molecular biology of secreted enzyme production by fungi. Crit. Rev. Biotechnol., 17: 273-306.
Direct Link - Argall, M.E., G.D. Smith, N.P.J. Stamford and B.N. Youens, 1992. Purification and properties of urease from the cyanobacterium Anabaena cylindrica. Biochem. Int., 27: 1027-1036.
Direct Link - Bermudez, E.M. and R.A. Burne, 1999. Genetic and physiologic characterization of urease of Actinomyces naeslundii. Inform. Immol., 67: 504-512.
Direct Link - Bongaerts, G.P.A., J. Uizetter, R. Brouns and G.D. Vogels, 1978. Uricase of Bacillus fastidiosus properties and regulation of synthesis. Biochimica Biophysica Acta (BBA)-Enzymol., 527: 348-358.
CrossRefDirect Link - Collier, J.L., B. Brahamsha and B. Palenik, 1999. The marine cyanobacterium Synechococcus sp. WH7805 requires urease (urea amidohydrolase, EC 3. 5. 1. 5) to utilize urea as a nitrogen source: Molecular-genetic and biochemical analysis of the enzyme. Microbiology, 145: 447-459.
Direct Link - Creaser, E.H. and R.L. Porter, 1985. The purification of urease from Aspergillus nidulans. Int. J. Biochem., 17: 1339-1341.
Direct Link - Farley, P.C. and S. Santosa, 2002. Reglulaiton of experssion of the Rhizopus oryzae uricase and urease enzymes. Can. J. Microbiol., 48: 1104-1108.
Direct Link - Fujinawa, S.B.G. and T.P. Dela, 1990. Application of acid urease to reduction of urea in commercial wines. Am. J. Enol. Viticult., 42: 350-354.
Direct Link - Ge, X., K. Cain and R. Hirschberg, 1990. Urea metabolism and urease regulation in the cyanobacterium Anabaena variabilis. Can. J. Microbiol., 36: 218-222.
Direct Link - Ghasemi, M.F., M.R. Bakhtiari, M. Fallahpour, A. Noohi, N. Moazami and Z. Amidi, 2004. Screening of urease production by Aspergillus niger strains. Iran. Biomed. J., 8: 47-50.
Direct Link - Hu, L.T., E.B. Nicholson, B.D. Jones, M.J. Lynch and H.L. Mobley, 1990. Morganella morganii urease purification, characterization and isolation of gene sequences. J. Bacteriol., 172: 3073-3080.
Direct Link - Jahns, T., U. Schafer and H. Kaltwasser, 1995. Heat-stable ureases from two Filamentous cyanobacteria. Microbiology, 141: 737-741.
Direct Link - Jahns, T., 1995. Purification and properties of urease from Sporobolomyces roseus. Antonie van Leeuwenhoek, 68: 209-214.
Direct Link - Jin, M., W. Rosario, E. Watler and D.H. Calhoun, 2004. Development of a large-scale HPLC-based purification for the urease from Staphylococcus leei and determination of subunit structure. Protein Exp. Purification, 34: 111-117.
Direct Link - Kakimoto, S.S.Y., S. Akiyama and Y. Nakao, 1989. Purification and characterization of acid urease from Lactobacillus reuteri. Agric. Biol. Chem., 53: 1119-1125.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lubbers, M.W., S.B. Rodriguez, N.K. Honey and R.J. Thornton, 1996. Purification and characterization of urease from Schizosaccharomyces pombe. Can. J. Microbiol., 42: 132-140.
Direct Link - Mackay, E.M. and J.A. Pateman, 1980. Nickel requirement of a urease-deficient mutant in Aspergillus nidulans. J. Gen. Microbiol., 116: 249-251.
Direct Link - Mackey, E.M. and J.A. Pateman, 1982. The regulation of urease activity in Aspergillus nidulans. Biochem. Gen., 20: 763-776.
Direct Link - Mehta, N., J.W. Olson and R.J. Maier, 2003. Characterization of Helicobacter pylori nickel metabolism accessory proteins needed for maturation of both urease and hydrogenase. J. Bacteriol., 3: 726-734.
Direct Link - Mobley, H.L. and R.P. Hausinger, 1989. Microbial ureases: Significance, regulation and molecular characterization. Microbiol. Rev., 53: 85-108.
Direct Link - Mobley, H.L.T., M.D. Island and R.P. Hausinger, 1995. Molecular biology of microbial ureases. Microbiol. Rev., 59: 451-480.
Direct Link - Mirbod, F., R.A. Schaller and T. Cole, 2002. Purification and characterization of urease isolated from the pathogenic fungus Coccidioides imitis. Med. Mycol., 40: 35-44.
Direct Link - Mulrooney, S.B., S.K. Ward and R.P. Hausinger, 2005. Purification and properties of the Klebsiella aerogenes UreE metal-binding domain, a functional metallochaperone of urease. J. Bacteriol., 10: 3581-3585.
PubMed - Omar, S.A. and M.H. Abd-Alla, 2000. Physiological aspects of fungi isolated from root nodules of faba bean (Vicia faba L.). Microbiol. Res., 154: 339-347.
Direct Link - Palinska, K.A., T. Jahns, R. Rippka and M.N. Tandeau de, 2000. Prochlorococcus marinus strain PCC 9511, a picoplanktonic cyanobacterium, synthesizes the smallest urease. Microbiology, 146: 3099-3107.
Direct Link - Pandey, P.C. and V. Pandey, 1991. Urease purification from the seeds of Cajanus cajan and its application in a biosensor construction. Applied Biochem. Biotechnol., 31: 247-251.
Direct Link - Park, L.S. and R.P. Hausinger, 1995. Evidence for the presence urease apoprotein complexes containing UreD, UreF and UreG in cells that are competent for in vivo enzyme activation. J. Bacteriol., 177: 1947-1951.
Direct Link - Pearson, M., L. Michel, R. Hausinger, P. Karplus, 1997. Structures of Cys 319 variants and acetohydroxamate inhibited Klebsiella aerogenes urease. Biochemistry, 36: 8164-8172.
Direct Link - Rai, A.K. and S. Singh, 1987. Urease of blue-green algae (cyanobacteria) Anabaena doliolum and Anacystis nidulans. Curr. Microbiol., 16: 113-117.
Direct Link - Rai, A.K., 1989. Purification and properties of urease from a cyanobacterium Anabaena doliolum. FEMS Microbiol. Lett., 61: 319-322.
Direct Link - Remaut, H.N. Safarov, S. Ciurli and J. Van Beeumen, 2001. Structural basis for Ni2+ transport and assembly of the urease active site by the metallochaperone UreE from Bacillus pasteurii. J. Biol. Chem., 52: 49365-49370.
Direct Link - Singh, H.N., S.N. Bagchi and R.K. Singh, 1983. L-Methionine-DL-sulfoximine resistant Het+ Nif+ and Het-Nif-strains of Nostoc muscorum assimilating methylamine as ammonium nitrogen source. FEMS Microbiol. Lett., 20: 31-34.
Direct Link - Singh, S., 1990. Regulation of urease activity in the cyanobacterium Anabaena doliolum. FEMS Microbiol. Lett., 67: 79-84.
Direct Link - Smith, P.T., A.K.J. Douglas and N. Goodman, 1993. Isolation and characterization of urease from Aspergillus niger. J. Gen. Microbiol., 139: 957-962.
Direct Link - Walfridsson, M., M. Anderlund, X. Bao and B. Hahn-Hagerdal, 1997. Expression of different levels of enzymes from the Pichia stipitis XYL1 and XYL2 genes in Saccharomyces cerevisiae and its effects on product formation during xylose utilization. Applied Microbiol. Biotechnol., 48: 218-224.
Direct Link - Weatherburn, M.W., 1967. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem., 39: 971-974.
CrossRefDirect Link - Wikipedia, 2006. Urea from Wikipedia, the free encyclopedia 14: GNU free documentation license. Text of the GNU free documentation license. Wikipedia is a Registered Trademark of the Wikimedia Foundation, Inc., Version 1.2, November 2002, Copyright (C) 2000,2001,2002 Free Software Foundation, USA., Boston, MA 02110-1301.
- Won, H.S. and B.J. Lee, 2004. Nickel-binding properties of the C-terminal tail peptide of Bacillus pasteurii UreE. J. Biochem., 136: 635-641.
Direct Link - Wray, W., T. Boulikas, V.P. Wray and R. Hancock, 1981. Silver staining of proteins in polyacrylamide gels. Anal. Biochem., 118: 197-203.
CrossRefDirect Link - Zambelli, B., M. Stola, F. Musiani, K. De Vriendt and B. Samyn et al., 2005. UreG, a chaperone in the urease assembly process, is an intrinsically unstructured GTPase that specifically binds Zn2+. J. Biol. Chem., 6: 4684-4695.
Direct Link