
F. Azam
Rhizobiology Laboratory, Soil Biology Division,
Nuclear Institute for Agriculture and Biology, Faisalabad, Pakistan







ABSTRACT
Studies conducted under controlled laboratory conditions using 14C showed a reduction in the rate of decomposition of plant residues with increased chemical complexity of the material; glucose being the most rapidly decomposed and least transformed into humic compounds. Lignin carbon was the most recalcitrant, but up to 50% was transformed into stable humus fractions. Rate and extent of immobilization and remineralization of N decreased with increased complexity of the plant residues/components; maximum being observed for glucose and minimum for lignins. Immobilization and remineralization turnover of N was used to determine dynamics of microbial biomass as well as to test assumptions used for its quantification with chloroform fumigation method. Chloroform fumigation was found to cause a substantial increase in the extractability and mineralization of non-biomass N. Quality of plant residues had a significant bearing on mineralization of N and its interaction with native soil N. Residues with narrow C/N ratio and high content of labile C had a positive effect on release of N from soil organic matter and its availability to crop plants. A substantial and real added nitrogen interaction was observed following application of fertilizer N (more for NH4 than NO3) and leguminous plant residues. In plant experiments, the interaction was exhibited by a substantial increase in root biomass especially under salinity stress. Higher amounts of N were released from leguminous plant residues in the presence of NH4 than NO3. Residues from cereal crops like rice and wheat had a negative effect on the plant availability of N from soil organic matter rather than from applied fertilizer. Studies under field conditions compared leguminous and non-leguminous crops for biomass accumulation and grain yield. When used as a green manuring crop, maize had an effect similar to recommended dose of fertilizer. Green manuring of cereal crop residues like wheat, avena and barley also had a positive effect on yield of wheat.
PDF Abstract XML References
How to cite this article
F. Azam, 2002. Studies on Organic Matter Dynamics and Nitrogen Availability Using 14C and 15N. Journal of Agronomy, 1: 20-24.
DOI: 10.3923/ja.2002.20.24
URL: https://scialert.net/abstract/?doi=ja.2002.20.24
DOI: 10.3923/ja.2002.20.24
URL: https://scialert.net/abstract/?doi=ja.2002.20.24
INTRODUCTION
Nitrogen (N) is the nutrient element that most often limits the crop productivity. Logically therefore the use of nitrogenous fertilizers has resulted in significant improvement in crop yields. According to statistics by FAO (2001), about 42 million tons of fertilizer N is being used annually on a global scale for the production of 3 major cereal crops i.e., wheat, rice, and maize (17, 9 and 16 million tons, respectively). However, the crop plants are able to use about 50% of the applied fertilizer N, while 25% is lost from the soil-plant system through leaching, volatilisation and denitrification, causing an annual financial loss of ca 3 billion US$. An increase of 1% in the fertilizer N use efficiency is estimated to save more than US$ 234 million (Raum and Johnson, 1999). In addition to economic considerations, the potential pollution caused by nitrogenous fertilizers has become a serious concern over the past few decades (Bouwman, 1996; Smith et al., 1997). One of the several ways to tackle these problems is to look for appropriate measures that will supplement N availability, enhance fertilizer uptake by plants and decrease the losses. Organic manures, especially the green manures, have traditionally been accepted and practiced not only as a source of supplemental N but also as soil conditioner.
Organic matter amendment has multifarious beneficial effects on soil fertility and productivity. Nutrient dynamics and availability, water economy of the crops and overall soil health depends to a considerable extent on the quality and quantity of organic matter reaching the soil. However, the intensity of these effects depends to a great degree on the chemical composition of the residues. It is quite well documented that plant residues rich in easily degradable constituents are rapidly oxidized, make a meager contribution to the stable humus fractions but facilitate rapid turnover (immobilization-remineralization) of nutrients (Azam et al., 1985; Haider and Azam, 1982; Kassim et al., 1981; Stott et al., 1983). On the contrary, residues high in lignins contribute substantially to the soil humus, are slow to decompose and thus generate a lower microbial activity. Plant residues with high N content and narrow C/N ratio (legumes) are reported to improve crop yields mainly by supplementing N supplies to plants. The release of N from organic N including plant residues could be enhanced by the application of nitrogenous fertilizers, the effect being more with NH4+ fertilizers (Azam et al., 1995; Hart et al., 1986; Woods et al., 1987). However, the net contribution of legume N to plants is generally low (Azam, 1990; Azam et al., 1985; Ladd et al., 1983) which suggests additional benefits in terms of overall improvement in soil fertility. It has also been observed that legume residues have a positive effect on the mineralization and plant availability of N from native soil organic matter through the so-called “priming effect” or “added nitrogen interaction” (Azam et al., 1995; Woods et al., 1987).
In plant experiments, the increase in biomass accumulation vis-a-vis enhanced N uptake is reported to result from increased root proliferation, which not only helps plants explore a greater soil volume for nutrient acquisition but is also useful in mitigating the negative effects of stresses like salinity and drought (Azam et al., 1992a,b). It is because of such effects of green manures and fertilizers that the plant uptake or mineralization from added source shows higher values when determined by “difference method” compared to isotopic method (Azam et al., 1993).
Organic manuring not only helps in economizing fertilizer N, but is more environment friendly and useful for a sustainable agricultural system. It is necessary, however, to develop a good understanding of the dynamics of organic matter and nitrogenous fertilizers, applied alone or in conjunction. Availability of stable and radioactive tracer techniques has been instrumental in achieving the required knowledge. From the Nuclear Institute for Agriculture and Biology (NIAB), Faisalabad, Pakistan, extensive studies have been reported on the dynamics of C and N (organic matter and nitrogenous fertilizers) and their implications to nutrient (especially N) cycling and crop production using stable and radioactive tracer techniques. This paper provides a summary of the information generated on I) basics of organic matter and N turnover in soil, ii) integrated use of nitrogenous fertilizers and plant residues, and iii) practical aspects of green manuring.
Materials and Methods
Transformation of plant residues and components: A Chernozem soil from Germany was amended with 1% uniformly 14C-labeled glucose (UL-glucose), cellulose (UL-cellulose), wheat straw (UL-straw) and corn stalk lignins specifically 14C-labelled at ring (L-Ring), side chain-2 (L-C2) or methoxyl (L-CH3). The amended soil was moistened to 60% water holding capacity with (15NH4)2SO4 and incubated for 12 weeks at 25 oC (Azam et al., 1985). During incubation, evolution of 14CO2 was measured on a scintillation counter. Soil samples were analyzed for total N and 15N using micro-Kjeldahl and mass spectrometric methods, respectively, to study the pattern of immobilization-remineralization as affected by different C sources. At the end of incubation, soil samples were also analyzed for the determination of 14C in humic acid, fulvic acid and humins using NaOH-Na4P2O7 as an extractant. Liquid samples were counted directly, while soil and humin samples were combusted before subjecting to liquid scintillation counting.
Mineralization of N from plant residues and soil in different soils: An experiment was conducted to study I) the mineralization of N from 15N-labelled soybean (Glycine max L.), vetch (Vicia villosa Roth.) tops and corn (Zea mays L.) stover in six Mollisols from Illinois, and ii) the interaction of applied residue N with the native soil N (Azam et al., 1993). Plants were grown in hydroponics containing 15N-labelled potassium nitrate. Harvested, dried and finely powdered plant residues were mixed with soil at 0.5% and incubation carried out at 25 oC. After 4 weeks of incubation, aliquots of freeze-dried soil were analyzed for mineral 15N and the release of residue N determined by isotope dilution equations. In a separate incubation study, the effect of NH4+-N and NO3- N on the mineralization of N from vetch was studied (Azam et al., 1995). 15N-labelled (3.3 atom % 15N) vetch was applied at 1% either with or without the addition of NH4+ and NO3- at two rates i.e., 100 and 200 mg kg-1 soil. Soil samples were analyzed for total N and 15N at different time intervals.
In a laboratory incubation experiment, effect of substrate addition rate and soil treatments extractability of newly immobilized 15N and native soil N was studied (Azam et al., 1989). The soil was amended with increasing levels of (15NH4)2SO4 and glucose (C/N ratio of the added material maintained at 30) to get added N concentration in soil of 67, 133, 200, 267 and 333 mg kg-1. The objective was to see if the amount of added C and N had an effect on extractability and hence plant availability of native soil N. At 106 hr after incubation (the time required for complete immobilization of applied N), aliquots of soil were subjected to CHCl3 fumigation (Jenkinson and Powlson, 1976). Portions of fumigated and unfumigated samples were extracted with 0.5M K2SO4 and the extracts subjected to analysis for total N and 15N.
Pot experiments on the response of plants to organic matter and nitrogen application: Several experiments were conducted to I) compare 5 rice varieties for exploiting soil N using 15N-labelled fertilizer (Azam et al., 1991a), ii) study the effect of wheat straw on the availability of soil-N and fertilizer-N to wetland soil (Azam et al., 1991b), and iii) to study the comparative effects of organic and inorganic nitrogen on rice yield and availability of N (Azam, 1990).
In experiment (I), seedlings of 5 rice cultivars were transplanted in potted soil fertilized with 15N-labelled ammonium sulphate. The plants were harvested at maturity and sources of plant N determined by isotopic analyses. The main objective was to study the influence of different plant types on the availability of native soil N.
In experiment (ii), rice nursery was transplanted in potted soil amended with 0, 0.1, 0.2 and 0.3% powdered wheat straw in combination with 0, 120, 240, and 360 mg kg-1 of (15NH4)2SO4-N. The objective was to study the interaction of organic matter and inorganic N in terms of availability of soil-N and fertilizer-N. The plants were grown under flooded conditions and harvested at maturity. The amount of soil-N and fertilizer-N in the whole plant was determined.
Experiment (iii) was aimed at differentiating between the sources of plant N. In this experiment, soil samples were treated with a) inorganic N as (15NH4)2SO4, b) 15N-labelled plant material of Sesbania aculeata, c) or the two sources together with change in 15N-labeling. Rice plants were grown to maturity and sub-samples analyzed for total N and 15N.
Field experiments on the use of green manures: Conventionally, leguminous green manures have been used, especially for rice in wheat-rice rotations prevalent in South Asian countries. In some exploratory field experiments, we have used fodder maize, avena, barley and wheat for green manuring. Multiple plots were prepared to allow for inclusion of different green manuring and fertilizer treatments. After 2 months of growth, maize plants were incorporated into the soil, with or without mineral fertilizer. For comparison, fertilizer treatment was also included. Two wheat varieties (Inqilab and WL-41) were tested for their response to different treatments. Wheat crop was harvested at maturity and data on grain yield and total biomass recorded.
RESULTS AND DISCUSSION
Decomposition of simple carbon compounds was rapid and a flush of decomposition was observed during the first few days (Fig. 1). Subsequent slow rate of CO2 evolution indicated mainly the decomposition of microbial products. Decomposition of lignin carbons, especially ring carbons was very slow and after 12 weeks of incubation an average of 22% of the lignin C was lost as CO2. Such differences have commonly been observed (Kassim et al., 1981a,b; Martin et al., 1971, 1980). Only a small proportion of glucose, cellulose and wheat straw was transformed into stable humus fractions, whereas a greater portion (>50%) of lignin carbons, especially from the ring, ended up in humic acid and fulvic acid (Fig. 2). Of the residual ring C, 56-67% was recovered in humic compounds, a greater proportion being in humic acid fraction. While transformation of C from simple compounds into humus fractions may result from synthetic activities of microorganisms, a significant proportion of lignin carbons could have been stabilized directly.
Differences in susceptibility of plant residues and components to microbial decomposition and transformation were also reflected in immobilization-remineralization of added 15N (Fig. 3; Azam et al., 1985). Glucose that is highly labile caused a rapid turnover of 15N, the process being slowed down with the increased complexity of the applied C. Similar results have been obtained by other workers (Ahmad et al., 1969; Marumoto et al., 1980). The variation in organic materials in terms of turnover of nutrients, especially N is of great significance for plant growth and productivity. This is one reason that organic materials high in N content and low in C/N ration are generally recommended for field application to maintain sufficient available N. However, as will be revealed later, the major effect of high C/N ration in plant residues is on blocking the release of N from soil rather than from applied fertilizer.
Chemistry of plant residues not only affects the immobilization- remineralization of N from other sources, but release of their own N is also significantly affected depending upon the nature of soil and the environmental conditions (Fox et al., 1990; Palm and Sanchez, 1991).
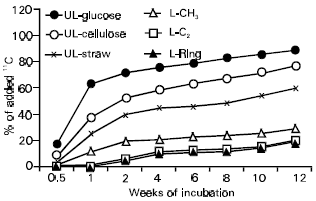 | |
| Fig. 1: | Decomposition (% of added 14C lost as CO2 from 14C-labelled plant residues and components. |
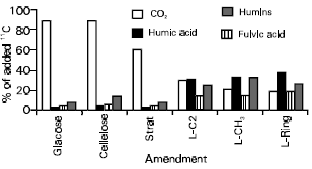 | |
| Fig. 2: | Transformation of 14C from different plant residues and components into CO2 humus fractions and biomass |
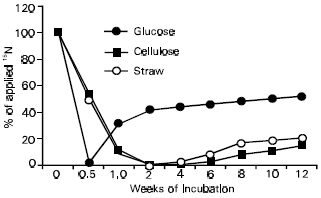 | |
| Fig. 3: | Immobilization and remineralization of applied 15N during the decomposition of glucose, cellulose and straw |
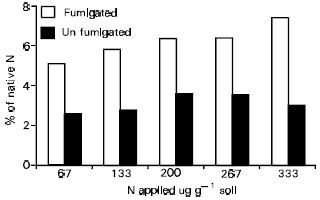 | |
| Fig. 4: | Effect of C and N addition of extractability of soil N from fumigated and unfumigated soil |
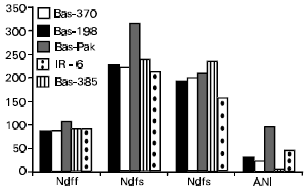 | |
| Fig. 5: | Uptake of soil N (Ndfs) from fertilized (second set of bars) and unfertilized (third set of bars) soil by five varieties of rice grown under flooded condition |
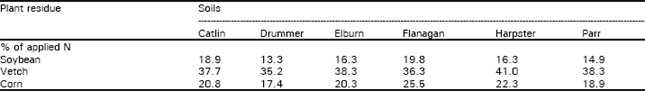
Results presented in Table 1 show that 13-41% of the residue N may be mineralized in a matter of few weeks. The rate and extent of mineralization depends not only on N content and C/N ratio but also on the proportion of labile C compounds. In this study, vetch which had higher N content as well as high proportion of labile C, was more quick to release N in mineral forms (for details, Azam et al., 1993). Release of N from corn was greater compared with soybean. The difference was attributed to N content of the two when grown under hydroponic conditions for 3 weeks. Further, rate of mineralization of N from plant residues can significantly be enhanced by the addition of mineral N, especially NH4+-N (Azam et al., 1995; Fog, 1990).
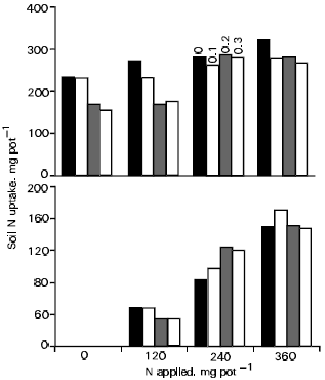 | |
| Fig. 6: | Effect of wheat straw (0,0.1,0.2 and 0.3%) and fertilizer N on uptake of soil N (top) and fertilizer N (bottom) by rice |
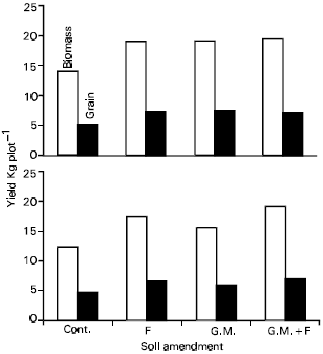 | |
| Fig. 7: | Grain and straw yield of two varieties of wheat i.e., Inqibal (top) and WL-1073 (bottom) as affected by fertilizer (F) application and/or green manure (G.M.) |
The addition of inorganic N not only enhances the mineralization of organic N, but has a positive effect on its extractability and thus potential availability to plants. In an study aimed at studying the effect of increasing levels of added C and N on the extractability of biomass and non-biomass N, it was found that extractability of newly immobilized N increased with the rate of amendment (Azam et al., 1989). Extractability was more from fumigated than unfumigated soil (Fig. 4) suggesting a higher susceptibility of microbial biomass to chemical extraction.
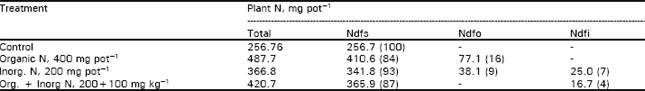
| Table 1: | Mineralization of N from 15N-labelled plant residues in different soils |
 | |
| Table 2: | Origin and amount of N in rice plants |
 | |
| Ndfs, N derived from soil; Ndfo, N derived from organic material; Ndfi, N derived from Inorganic fertilizer | |
These results also revealed that non-biomass N is also subject to extraction and inclusion with the biomass thereby giving inflated values for the biomass N. In addition, the extractability increased with the amount of amendment, an observation supported by other studies (Hart et al., 1986).
By releasing carbonaceous materials into the soil, the plants also influence the release of N from soil organic matter, the so-called root-induced N mineralization (Breland and Bakken, 1991; Hart et al., 1979). The extent of release was different with the type of plant ( Fig. 5). In this study, 5 rice varieties were grown in unfertilized or fertilized (15N-labelled ammonium sulphate) soil and at harvest data on N derived from soil and the resulted added nitrogen interaction (ANI) was computed. The varieties showed substantial variation in uptake of N from soil, whereas fertilizer N uptake was not so different. Fertilizer application did have a positive effect on uptake of soil N leading to ANI the extent of which was different in the varieties. This difference could be attributed to the rooting characteristics as well as response to fertilizer application. In another study, root proliferation had a significant bearing on uptake of N from soil organic matter as well as tolerance of plants to salinity stress (Azam et al., 1992a,b). This objective could be achieved by increasing the amount of fertilizer N applied. These studies suggested that an increase in root proliferation could help plants withstand stresses like salinity and drought. To achieve this objective, an ambitious programme is underway on root modifications using microorganisms and growth hormones (Gulnaz et al., 1999).
Release in soil of carbonaceous compounds or application of plant residues may cause short-term or long-term immobilization of essential nutrients like N (Gok and Ottow, 1988). Hence, residues having wider C/N ratio are generally considered to have a negative effect on plant productivity and rightly so. Since, soil organic matter is the predominant source of plant N, any change in N availability from it will have a bearing on plant growth. This is one reason that there is a good relationship between availability of soil N and plant growth. In one of the studies, we observed that compared to fertilizer N, soil N uptake is more severely affected by plant residues like wheat straw (Fig. 6; Azam et al., 1991b). However, increase in the amount of fertilizer N applied was found to help in mitigating the negative effects of straw amendment, particularly its impact on uptake of soil N.
Compared to high C/N ratio cereal residues, those rich in N (leguminous) and having narrow C/N ratio not only provide N directly to the plants, but have a positive effect on the mineralization and plant availability of N from soil organic matter. Indeed, one of the benefits of green manuring may be seen in terms of enhanced N supplies indirectly through the “added nitrogen interaction”. A significant increase in soil N uptake by rice plants was indeed observed following application of legume residues (Table 2). In this case, addition of organic matter resulted in an uptake of 410.6 mg N from soil compared to 256.7 mg unamended soil (control). This could be expected and the results presented above support the notion that easily decomposable organic matter will enhance soil N mineralization as a result of overall increase in microbial activities. However, this kind of interpretation is seldom found in literature.
From the above it can be inferred that easily decomposable plant residues will have a bearing on availability of N from sources other than fertilizer and applied organic matter. Thus not only leguminous but non-leguminous residues as well having a good component of easily decomposable C may help crops perform better. For example, maize plants harvested at 6-8 weeks not only having an N content of 2-3%, but also contains 50% of the carbon in labile forms (Azam et al., 1993). Incorporation of such residues is expected to supplement N supplies both directly (through mineralization) as well as indirectly (through ANI). In a field experiment, such an effect was indeed demonstrated (Fig. 7). In this case, incorporation of maize residues had an effect similar to recommended rate of chemical fertilizer on two wheat varieties. Both grain and straw yield were positively affected, although varietal differences were observed.
In summary, the information provided above suggests that with proper management of plant residues, a sustainable increase in agricultural productivity could be achieved. Care needs to be taken that nutrient dynamics are not affected negatively by residue incorporation.
REFERENCES
- Ahmad, Z., H. Kai and T. Harada, 1969. Factors affecting immobilization and release of N in soil and chemical characteristics of N newly immobilized. II. Effect of carbon sources on immobilization and release of N in soil. Soil Sci. Plant Nutr., 15: 252-258.
PubMed - Azam, F., 1990. Comparative effects of organic and inorganic nitrogen sources applied to a flooded soil on rice yield and availability of N. Plant Soil, 125: 255-262.
CrossRefDirect Link - Azam, F., M. Ashraf, A. Lodhi and M.I. Sajjad, 1992. Fate and interaction with native soil N of ammonium N applied to wetland rice (Oryza sativa L.) grown under saline and non-saline conditions. Biol. Fertil. Soils, 13: 102-107.
CrossRefDirect Link - Azam, F., M. Ashraf, A. Lodhi and M.I. Sajjad, 1991. Relative significance of soil and nitrogenous fertilizer in nitrogen nutrition and growth of wetland rice (Oryza sativa L.). Biol. Fertil. Soils, 11: 57-61.
CrossRefDirect Link - Azam, F., A. Lodhi and M. Ashraf, 1991. Availability of soil and fertilizer nitrogen to wetland rice following wheat straw amendment. Biol. Fert. Soils, 11: 97-100.
CrossRefPubMedDirect Link - Azam, F., F.W. Simmons and R.L. Mulvancy, 1993. Mineralization of N from plant residues and its interaction with native soil N. Soil Biol. Biochem., 25: 1787-1792.
Direct Link - Azam, F., R.L. Mulvaney and F.W. Simmons, 1995. Effects of ammonium and nitrate on mineralization of nitrogen from leguminous residues. Biol. Fertil. Soils, 20: 49-52.
CrossRefDirect Link - Bouwman, A.F., 1996. Direct emission of nitrous oxide from agricultural soils. Nutr. Cycl. Agroecosyst., 46: 53-70.
CrossRefDirect Link - Breland, T.A. and L.R. Bakken, 1991. Microbial growth and nitrogen immobilization in the root zone of barley (Hordeum vulgare L.), Italian ryegrass (Lolium multiflorum Lam.) and white clover (Trifolium repens L.). Biol. Fertil. Soils, 12: 154-160.
Direct Link - Fox, R.H., R.J.K. Myers and I. Vallis, 1990. The nitrogen mineralization rate of legume residues in soil as influenced by their polyphenol, lignin and nitrogen contents. Plant Soil, 129: 251-259.
Direct Link - Gok, M. and J.C.G. Ottow, 1988. Effect of cellulose and straw incorporation in soil on total denitrification and nitrogen immobilization at initially anaerobic and permanent anaerobic conditions. Biol. Fert. Soils, 5: 317-322.
CrossRefPubMedDirect Link - Gulnaz, A., J. Iqbal, S. Farooq and F. Azam, 1999. Seed treatment with phytohormones and crop productivity. I. 2,4-D as an inducer of salinity-tolerance in wheat (Triticum aestivum L.). Plant Soil, 210: 209-218.
Direct Link - Hart, P.B.S., J.H. Rayner and D.S. Jenkinson, 1986. Influence of pool substitution on the interpretation of fertilizer experiments with 15N. Eur. J. Soil Sci., 37: 389-403.
CrossRefPubMedDirect Link - Jenkinson, D.S. and D.S. Powlson, 1976. The effects of biocidal treatments on metabolism in soil-V: A method for measuring soil biomass. Soil Biol. Biochem., 8: 209-213.
CrossRefDirect Link - Kassim, G., J.P. Martin and K. Haider, 1992. Incorporation of a wide variety of organic substrate carbons into soil biomass as estimated by fumigation procedure. Soil Sci. Soc. Am. J., 45: 1106-1112.
Direct Link - Martin, J.P., K. Haider and G. Kassim, 1980. Biodegradation and stabilization after 2 years of specific crops, lignins, and polysaccharide carbons in soils. Soil Sci. Soc. Am. J., 4: 1250-1255.
Direct Link - Palm, C.A. and P.A. Sanchez, 1991. Nitrogen release from some tropical legumes as affected by lignin and polyphenol contents. Soil Biol. Biochem., 23: 83-88.
PubMedDirect Link - Raun, W.R. and G.V. Johnson, 1999. Improving nitrogen use efficiency for cereal production. Agron. J., 91: 357-363.
CrossRefPubMedDirect Link - Stott, D.E., G. Kassim, W.M. Jarrell, J.P. Martin and K. Haider, 1983. Stabilization and incorporation into biomass of specific plant carbons during biodegradation in soil. Plant Soil J., 70: 15-26.
CrossRefPubMedDirect Link - Woods, L.E., C.V. Cole, L.K. Porter and D.C. Coleman, 1987. Transformations of added and indigenous nitrogen in gnotobiotic soil: A comment on the priming effect. Soil Biol. Biochem., 19: 673-678.
PubMedDirect Link