
M. Annamalai
Center for Biotechnology, Anna University, Chennai, India
Mukesh Doble
Department of Biotechnology, 11T Madras, Chennai, India
Journal of Applied Sciences
Year: 2007 | Volume: 7 | Issue: 15 | Page No.: 2051-2056







ABSTRACT
Mathematical modeling of hydantoinase production system from microbial sources, which would help to understand the mechanism of the process, has not been attempted earlier. This paper tries to model five state variables (biomass, substrate, product (D-hydantoinase), Oxygen Uptake Rate (OUR) and carbon dioxide production rate (CPR)) for three carbon sources namely glucose, glycerol and maltose in the production of D-hydantoinase using Agrobacterium radiobacter as source. Several models were tested to fit the aerobic batch experimental data from a 3 L bioreactor. The best fitting model consisted of (a) biomass growth non-linearly dependent on substrate concentration, (b) product formation rate following exponential form of product inhibition and (c) OUR following positive regulation by substrate. D-hydantoinase production in maltose experiences minimal lag phase and stronger product inhibition when compared to glycerol. Maltose showed higher biomass yield (0.25) and specific D-hydantoinase production (27.44 U mg-1) compared to glycerol whose values are 0.18 and 21.97 U mg-1, respectively.
PDF Abstract XML References Citation
How to cite this article
M. Annamalai and Mukesh Doble, 2007. Modeling of D-Hydantoinase Production by Agrobacterium radiobacter in a Batch System. Journal of Applied Sciences, 7: 2051-2056.
DOI: 10.3923/jas.2007.2051.2056
URL: https://scialert.net/abstract/?doi=jas.2007.2051.2056
DOI: 10.3923/jas.2007.2051.2056
URL: https://scialert.net/abstract/?doi=jas.2007.2051.2056
INTRODUCTION
D-Hydantoinase (E.C.3.5.2.2) is a family of enzymes that catalyses reversible hydrolytic ring cleavage of hydantoins or 5-substituted hydantoin derivative, which includes alkyl or aryl-substituted hydantoins like phenyl hydantoin, D-hydroxy phenyl hydantoin etc. It is an industrially valuable enzyme for the synthesis of D-amino acids like D-p-hydroxyphenylglycine, D-valine, D-phenylglycine and D-serine (Achary et al., 1997; Lee et al., 1997). Mathematical modeling of the fermentation process helps to elucidate the mechanism of production process; understand the nature of inhibition exerted on the growth of microorganism, estimate kinetic parameters such as specific growth rate of biomass and product formation rate, develop understanding between effects of operational conditions on production and reduce laboratory experiments thereby saving time and resources. Literature related to hydantoinase modeling to the best of the authors knowledge is non-existent but there have been other efforts, such as streptokinase system (Patnaik, 1995); lactic acid fermentation (Biazar et al., 2003) and oxygen demand in amino acid production (Yao et al., 2001).
This study tries to obtain a mathematical model from batch fermentation data for D-hydantoinase production that will predict the time profile of five state variables namely biomass production, substrate consumption, product formation, OUR and CPR for three different carbon sources as substrates namely glucose, maltose and glycerol.
EXPERIMENTAL METHODOLOGY
The batch experimental data for D-hydantoinase production (consisting of the time profile of state variables namely, biomass, substrate and product, OUR and CPR for maltose, glycerol and glucose) used here for modeling was obtained from the laboratory experiments conducted earlier (Achary, 1997). The production of D-hydantoinase was done with Agrobacterium radiobacter NRRL B 11291. The experiments were performed in a 3 L cylindrical reactor with a working volume of 2 L operated with a mechanical agitator. Since the process is aerobic, dissolved oxygen concentration was maintained at high levels by sparging air at high rate. Biomass was measured off-line sphectrophotometrically by absorbance at the rate of 600 nm. Paramagnetic and infrared exhaust gas analyzers were used to determine the levels of oxygen and carbon dioxide in the exhaust gas, respectively.
MODELING METHODOLOGY
The general equation for rate of biomass (X) production, substrate (S) consumption, product (P) formation, OUR and CPR are given by the following set of general differential Eq. 1-7.
| (1) |
| (2) |
| (3) |
| (4) |
| (5) |
| (6) |
| (7) |
where:
| μ | = | The specific growth rate (h–1). |
| qs | = | The specific rate of substrate consumption (g L–1h–1OD–1). |
| qp | = | The specific rate of product formation (U mL–1h–1OD–1). |
| qo | = | The specific rate of oxygen uptake rate (mM h–1 OD–1). |
| = | The specific rate of carbon dioxide production (mM h–1OD–1). | |
| Cl | = | The dissolved oxygen concentration (mM). |
| kla | = | The volumetric oxygen transfer coefficient (h–1) = 227.47 h–1. |
| C* | = | Saturation level of dissolved oxygen concentration (0.2325 mM). |
The differential equations were integrated using ODE solvers available in MATLAB with appropriate initial conditions for the state variables while the parameters in the model were estimated by fitting the model predictions with experimental data. Sum of square of errors (SSE) was chosen as the objective function to serve as criteria to judge the predictive capability of the model, which was minimized. The minimization was performed using Newton’s least square method, Simplex method and Constrained minimization method available in MATLAB.
RESULTS AND DISCUSSIONS
The following are some of the important inferences drawn on the mechanism of D-hydantoinase production based on the plots of experimental data in Fig. 1a and b.
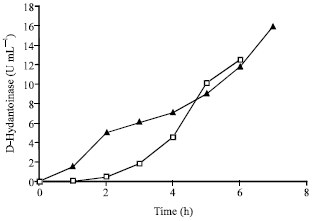 | |
| Fig. 1a: | Experimental product amount vs. time for carbon sources maltose |
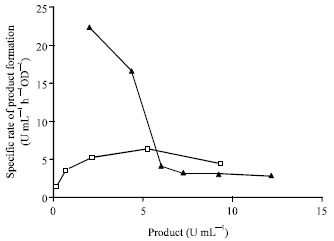 | |
| Fig. 1b: | Experimental Specific rate of product formation vs. product amount for carbon sources maltose |
Figure 1a suggests a considerable lag in product formation in the case of glycerol while it is minimum in maltose.
The rate of product formation is found not to be proportional to growth rate but depends upon the accumulation of biomass and the specific product yield from biomass. (Braun and Sutherland, 2003) also observed that most enzyme production occurred during the transition from exponential to stationary phase. There has also been usage of different models for growth and product formation rates (Yao et al., 2001) for amino acid production, again indicating lack of proportionality between the above two rates. Hence it would be suitable to use different models for biomass growth and product formation rate. Figure 1b shows that product formation in case of maltose is significantly inhibited by hydantoinase while product inhibition is weak in the case of glycerol.
D-Hydantoinase production in batch experiment with glucose is negligible due to catabolite repression (Lee et al., 1997) and hence product formation for this carbon source is hence forth not discussed. OUR was growth associated for maltose and glycerol, as observed in clavulanic acid production (Baptista-Neto et al., 2000) and it is higher for former when compared to latter because of positive regulation by substrate.
Nearly twenty six models, consisting of several simple (Monod, classical form of substrate and/or product inhibition, Contois, Teisser and Mosser for growth rate, classical form of substrate and/or product inhibition, growth and non growth associated models for rate of product formation), non-linear models (growth rate with non linear dependence on substrate (Yao et al., 2001)), logarithmic model of product formation (Yao et al., 2001), exponential form of product and/or substrate inhibition (Biazar et al., 2003) and few linear models of substrate and /or product inhibition (Xiu et al., 2004), were tried to fit the batch experimental plots of X, S, P. Various models for OUR were attempted which includes Monod, growth associated and non-growth associated oxygen uptake, simple and exponential form of substrate and oxygen inhibition and exponential form of positive substrate regulation model. The models tried for CPR includes growth and non growth associated models. The best model which gave the lowest combined SSE (6.82) and R2 = 0.985 was:
| (11) |
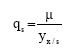 | (12) |
| (13) |
| (14) |
| (15) |
The best model for μ for all the three substrates was the non-linear model (Yao et al., 2001). In accordance to the conclusions made earlier for product formation rate from Fig. 1a and b, qp in the best model includes product inhibition term in exponential form as in e (-(P/k1)n1) while the lag in product formation at high substrate concentration is expressed by the term e (-(S/k2)n2), which can be used to explain the lag phase by imposing negative constraint on n2.
| Table 1: | Parameters estimated for the best models |
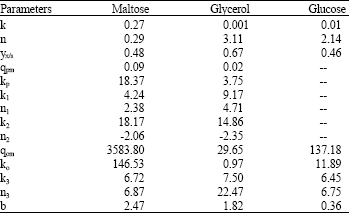 | |
In Eq. 11-14, k is specific rate at which substrate is used for growth, yx/s is specific yield of biomass (OD L g–1), qpm is maximum specific product formation rate (U mL–1 h–1OD–1), qom is maximum specific oxygen consumption rate (mM h–1OD–1), ko is the Monod saturation constant in qo (mM), kp is the Monod saturation constant in qp (g L–1), k1 is the product concentration at which product inhibition term e (-(P/k1)n1) is 0.368, k2 is the substrate concentration at which the lag phase term in qp, e (-(S/k2)n2) is 2.72, k3 is the substrate concentration at which positive regulation by substrate term in qo, e (-(S/k3)n3) is 2.72, b, n, n1, n2 and n3 are constants. The corresponding parameters for the three carbon sources for the best model are included in Table 1.
The comparison of the best model prediction to the experimental data for substrate and product formation is shown in Fig. 2, for biomass formation in Fig. 3 and for OUR and CPR in Fig. 4 for maltose. Similar figures for other substrates are not shown.
The specific growth rate μ in the best model is represented by k.Sn, where k and n describe the growth of Agrobacterium in different substrates. Higher k and lower n values mean that the biomass growth is balanced, less dependent on the external conditions such as availability of substrate and a longer exponential phase (Yao et al., 2001). Glycerol seems to possess the highest n and lowest k, due to which it supports highly uneven growth as described above, while maltose provides balanced growth and is least affected by the substrate concentration (it has a lower n value). Glycerol is transported by facilitated diffusion (Schweizer et al., 1997) due to which it equalizes its own concentration inside and outside the cell. Hence growth rate would be strongly dependent on substrate concentration. This is indicated by high value of n in the growth rate equation in the present study. The Maltose transporter is an ATP dependent ABC transporter (Boos and Shuman, 1998) which is specific with high affinity among the three substrate transporter. The rate of maltose transport by ATP dependent permease follows Michaelis Menten form (Nelson and Cox, 2003). The low saturation constant (in the Michelis Menten equation) leads to the rate equation independent of concentration, which is analogous to a low n in the growth rate equation in the best model.
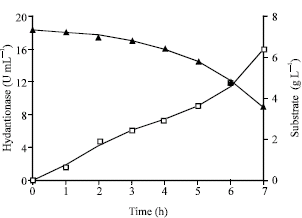 | |
| Fig. 2: | Comparison of model prediction with experiment for substrate consumption and D-hydantoinase production for maltose in batch cultivation ( |
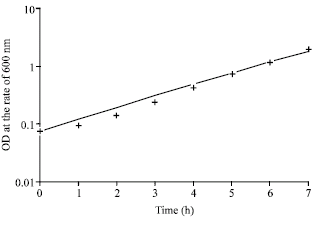 | |
| Fig. 3: | Comparison of best model prediction with experiment for biomass production for maltose. (+ = experimental data while continuous line = model) |
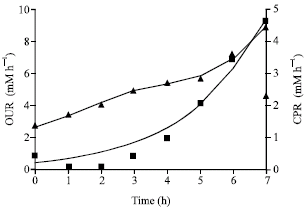 | |
| Fig. 4: | Time profile of OUR and CPR for maltose in batch cultivation for D-hydantoinase production. Markers ( |
Glucose and glycerol have comparable n and they support similar growth. (Yao et al., 2001) reported values of k and n to be 0.00173 and 0.776, respectively for L-lysine batch fermentation by Brevibacteria lactofermentum with glucose as substrate for dissolved oxygen level of 20%. In this study, k and n for glucose are 0.01 and 2.14, respectively for Agrobacterium radiobacter.
The specific biomass yield (yx/s) obtained from the best model can be converted to dimensionless yield (yx/s)a using the conversion factor 1 OD = 0.38 g L–1 (Lee et al., 1997). The dimensionless yield (yx/s)a for glucose, maltose and glycerol are 0.175, 0.18 and 0.25, respectively for Agrobactrium radiobacter. (yx/s)a was reported to be 0.15 for streptokinase production (Patnaik, 1995) and 0.16 for D-hydantoinase production in batch cultivation using glucose as carbon source (Lee et al., 1997). The latter value is closer to model value of (yx/s)a estimated for glucose in this study. There seems to be no difference between yield of glucose and maltose. Glycerol is a non-fermentable carbon source as a result of which yield of by-products like lactic acid, ethanol etc. is nil resulting in highest biomass yield. The specific product yield (yp/x)a found by our modeling study is 27.44 U mg–1 for maltose and 21.97 U mg–1 for glycerol which matches with the literature finding for Agrobacterium radiobacter of 25 U mg–1 for maltose and 14 U mg–1 for glycerol (Achary et al., 1997) and a value of 25 U mg–1 for fed-batch cultivation of recombinant E. coli harboring the plasmid for D-hydantoinase expression using glucose (Lee et al., 1997).
It was observed that D-hydantoinase production in maltose experiences more product inhibition than in glycerol (Fig. 1b) which is also seen from the fact that k1 in the model is lower for maltose than that for glycerol. Lactic acid inhibition has been included in streptokinase production (Patnaik, 1995). So the possible inhibition by lactic acid or other acids in glycerol is very less as it is non fermentable and hence the total acid yield from it is much lesser than maltose. It was reported by (Lucero et al., 2000) that Saccharomyces cerevisiae accumulates trehalose more when grown in non-fermentable sugars like glycerol compared to sugars like maltose. Trehalose accumulation reduces the inhibition on product formation by sugar phosphate (Bonini et al., 2000). Since sugar phosphate accumulation is lower in glycerol than maltose due to increased trehalose accumulation, product inhibition in the case of former is less than in latter. It can be inferred from the negative n2 in the best model for both maltose and glycerol that product formation in both the substrates faces lag at high substrate concentration, confirming the interpretations made graphically earlier based on Fig. 1a. However, maltose causes minimum lag phase when compared to glycerol which can be inferred from higher k2 in the best model for the former than for the latter. Xiu et al. (2004) in their clauvulanic acid production process, when Streptomyces clavuligerus NRRL 3585 was cultivated with glycerol as carbon and energy source, a lag period of nearly nine hours was observed. Clauvulanic acid is a strong β-lactamase inhibitor, a secondary metabolite that is produced in the late logarithmic phase. Clauvulanic acid is similar to D-hydantoinase production since in both cases product formation and biomass growth rate are non-proportional. It was also remarked by them that maltose was easily metabolized by Streptomyces clavuligerus which implies that it would support faster and steady growth rate than glycerol leading to faster accumulation of biomass. It was reported by Achary et al. (1997) that D-hydantoinase batch production was higher in maltose (22.5 U mL–1) compared to glycerol (17.5 U mL–1) using Agrobacterium radiobacter. This view that overall maltose yields overall higher volumetric production is supported by the fact higher the k and lower the n in μ would ensure sufficient biomass to accumulate by maintaining a steady growth rate and also would direct more carbon source to product formation (Yao et al., 2001). The specific product yield, (yp/x)a and qpm is also higher for maltose than glycerol. Overall OUR and CPR (b is higher for maltose) is also higher for maltose than glycerol.
CONCLUSIONS
Several models ranging from simple classical to mathematically complicated functions describing the substrate and product inhibitions were tested to fit batch experimental data for D-hydantoinase production using three different substrates namely maltose, glycerol and glucose. The best model showed non-linear dependence of specific growth rate (μ) on substrate, specific substrate consumption rate (qs) was proportional to μ and specific product formation rate (qp) obeyed exponential form of product inhibition. Product formation was preceded by lag phase. Growth rates with glycerol and glucose were strongly dependent on substrate concentration while steady growth was observed with maltose. D-Hydantoinase production in maltose experienced higher product inhibition and minimal lag phase when compared with maltose. Final production (both volumetric and specific) of D-hydantoinase was higher with maltose (27.44 U mg–1) than with glycerol (21.9 U mg–1). Oxygen uptake rate was positively regulated by substrate while carbon dioxide production rate seemed to be non growth associated. Positive regulation by the substrate on OUR was higher in maltose when compared to glycerol. Both oxygen uptake and carbon dioxide production rates were higher in maltose than in glycerol. A good model can be used to plan the operational strategy to achieve the required target; extend it to a continuous process mode and can be used to optimize operational parameters like flow rate, substrate concentration in feed etc. to achieve maximum productivity and can also be used to study the effect of change of operational parameters on the stability of the system and identify non-linear phenomenon.
REFERENCES
- Achary, A., K.A. Hariharan, S. Bandhyopadhyaya, R. Ramachandaran and K. Jayaraman, 1997. Application of numerical modeling for the development of optimized complex medium for D-hydantoinase production from Agrobacterium radiobacter NRRL B 11291. Biotechnol. Bioeng., 55: 148-154.
Direct Link - Baptista-Neto, A., E.R. Gouveia, A.C. Jr. Badino and C.O. Hokka, 2000. Phenomenological model of the clavulanic acid production process utilizing Streptomyces Clavuligerus. Braz. J. Chem. Eng., 17: 4-7.
CrossRefDirect Link - Biazar, J., M. Tango, E. Babolian and R. Islam, 2003. Solution of the kinetic modeling of lactic acid fermentation using Adomian decomposition method. Applied Math. Comput., 144: 433-439.
CrossRefDirect Link - Bonini, B.M., C. Van Vaeck, C. Larsson, L. Gustafsson, P. Ma, J. Winderickx, P. Van Dijck and J.M. Thevelein, 2000. Expression of Escherichia coli otsA in a Saccharomyces cerevisiae tps1 mutant restores trehalose 6-phosphate levels and partly restores growth and fermentation with glucose and control of glucose influx into glycolysis. Biochem. J., 350: 261-268.
PubMedDirect Link - Boos, W. and H. Shuman, 1998. Maltose/maltodextrin system of Escherichia coli: Transport, metabolism and regulation. Microbiol. Mol. Biol. Rev., 62: 204-229.
Direct Link - Braun, P. and J.P. Sutherland, 2003. Predictive modeling of growth and enzyme production and activity by a cocktail of Pseudomonas sp., Shewanella putrefaciens and Acinetobacter sp. Int. J. Food Microbiol., 86: 271-282.
Direct Link - Lee, D.C., G.J. Kim, Y.K. Cha, C.Y. Lee and H.S. Kim, 1997. Mass production of thermostable D-hydantoinase by batch culture of recombinant Escherichia coli with a constitutive expression system. Biotechnol. Bioeng., 56: 449-455.
Direct Link - Lucero, P., E. Penalver, E. Moreno and R. Lagunas, 2000. Internal trehalose protects endocytosis from inhibition by ethanol in Saccharomyces cerevisiae. Applied Environ. Microbiol., 66: 4456-4461.
Direct Link - Patnaik, P.R., 1995. A heuristic approach to fed-batch optimization of streptokinase fermentation. Bioprocess Eng., 13: 109-112.
CrossRefDirect Link - Schweizer, H.P., R. Jump and C. Po, 1997. Structure and gene-polypeptide relationships of the region encoding glycerol diffusion facilitator (glpF) and glycerol kinase (glpK) of Pseudomonas aeruginosa. Microbiology, 143: 1287-1297.
CrossRefDirect Link - Yao, H.M., Y.C. Tian, M.O. Tade and H.M. Ang, 2001. Variation and modeling of oxygen demand of oxygen demand in amino acid production. Chem. Eng. Processing, 40: 401-409.
CrossRefDirect Link