
Muhammad Jamil
Eui Shik Rha
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 4 | Page No.: 654-658







ABSTRACT
The study was carried out to assess whether water uptake could be improved in sugar beet seeds and salt tolerance at the germination and early seedling stage by soaking the seeds for 10 h in distilled water (control), 100, 150 and 200 mg L-1 GA3. Electrical Conductivity (EC) values of the NaCl solution were 0.0 (control), 4.7, 9.4 and 14.1 dS m-1 NaCl. Priming increased the final germination percentage and the germination rate (1/t 50, where t 50 is the time to 50% of germination) under saline condition. Water uptake of primed seeds also increased significantly with increasing concentration of GA3 as compared to control. Priming also alleviated the adverse effect of salt stress on sugar beet in terms of roots and shoots lengths and fresh weights of plants, roots and shoots.
PDF Abstract XML References Citation
How to cite this article
Muhammad Jamil and Eui Shik Rha, 2007. Gibberellic Acid (GA3) Enhance Seed Water Uptake, Germination and Early Seedling Growth in Sugar Beet under Salt Stress. Pakistan Journal of Biological Sciences, 10: 654-658.
DOI: 10.3923/pjbs.2007.654.658
URL: https://scialert.net/abstract/?doi=pjbs.2007.654.658
DOI: 10.3923/pjbs.2007.654.658
URL: https://scialert.net/abstract/?doi=pjbs.2007.654.658
INTRODUCTION
Salinity is one of the major problem to increasing production in crop growing areas throughout the world. Numerous attempts have been made to improve the salt tolerance of crops by traditional breeding programmes, but commercial success has been very limited (Santa-Cruz et al., 2002). Pre-sowing seed treatment or seed priming is an easy technique and an alternative approach recently used to overcome salinity problems. Priming (osmoconditioning) is one of the physiological methods, which improves seed performance and provides faster and synchronized germination (Sivritepe and Dourado, 1995). Pre-soaking or priming seeds of a number of crops has improved germination, seedling establishment and, in some cases, stimulated vegetative growth and hence crop yield (Amzallag et al., 19901; Kaur et al., 1998).
Application of abiotic stresses results in an altered levels of plant growth hormones and decreased plant growth (Morgan, 1990). Plant Growth Regulators (PGRs) have been found to play a central role in the integration of the responses expressed by plants under stress conditions (Amzallag et al., 1990). PGRs may also enhance germination and adaptation of plants to stress conditions (Banyal and Rai, 1983). Gibberellic acid and kinetin have been reported to increase percentage germination and seedling growth (Kaur et al., 1998).
Seed priming has been successfully demonstrated to improve germination and emergence in seeds of many crops, particularly seeds of vegetables (Heydecker and Coolbaer, 1977). The beneficial effects of priming have also been demonstrated for many field crops such as sugar beet, wheat, maize, soybean and sunflower (Singh, 1995; Parera and Cantliffe, 1994; Khajeh-Hosseini et al., 2003; Sadeghian and Yavari, 2004).
The objective of this study was to assess the effect of different seed treatment (100,150 and 200 mg L-1) with GA3 on germination and early seedling growth of sugar beet under salt stress conditions and whether priming improve water uptake in sugar beet seeds.
MATERIALS AND METHODS
Seeds of sugar beet (Beta vulgaris L. cv. Tianjin qing pielan) were obtained from Institute of Soils and Fertilizers, Beijing, China. Seeds were surface sterilized in 10% sodium hypochlorite solution for 10 min, then rinsed with sterilized distilled water and air-dried at an ambient temperature of 25°C in the laboratory. The sterilized seeds were soaked for 10 h in 100, 150 and 200 mg L-1 GA3 or distilled water. After the pre-sowing treatment, all seed samples were rinsed with distilled water and dried in an oven at 30°C for two hour to eliminate surface moisture.
Seeds were germinated in sterilized disposable Petri dishes (87 mm diameter, 15 mm height) containing two Whatman No. 2 filter papers soaked with 10 mL of distilled water or 4.7, 9.4 and 14.1 dS m-1 NaCl concentrations. Petri dishes were sealed with parafilm to prevent evaporation of water, thus minimizing the changes in concentration of the solutions. Seeds were incubated in growth chamber at 25°C. Five replicate dishes with 20 seeds per replicate were used for each treatment. Seeds were hand sorted to eliminate broken and small seeds. Seed germination was evaluated after every 12 h up to 8 days and germination rate was calculated as 1/t50 (where t50 is the time to 50% of germination) (Jamil and Rha, 2006). After 36 h, seeds started to germinate (seeds were considered to be germinated with the emergence of the radicle). In all treatments a continuous increase in the number of germinating seeds as well as in the lengths of roots and shoots was observed during the frequent days of germination. The lengths of roots and shoots of the germinated seeds, which were more than 2 mm in length, were measured and recorded after 15 days of sowing. The fresh weights of plants, roots and shoots were also determined after 15 days.
Two grams of the seeds from each seed treatment were placed in petri dishes containing 20 mL distilled water for 2 h to determine water uptake of seeds. The water uptake was expressed as the percentage increase in moisture content on fresh weight basis.
The experiment was designed by using a randomized complete block design with three replications. Analysis of variance was performed by using the Microsoft Excel version 5.0 (Middleton, 1995). Means values for different plant characteristics were compared through LSD test (Li, 1964).
RESULTS
Salinity caused a significant reduction in germination percentage with increasing salt stress (Fig. 2A). There was considerable reduction in germination at highest level of salinity (9.4 and 14.1 dS m-1 NaCl). However, priming increased germination percentage under salt stress (Fig. 2A). Of all priming treatments, 150 and 200 mg L-1 of GA3 was the most effective in increasing germination under salt stress. Water uptake of primed seeds also increased significantly with increasing concentration of GA3 as compared to control (Fig. 1).
The final germination rate of control and primed seeds under various conditions of salinity was expressed as a 1/t50 of the germination of seeds of the same population as in control. The germination response of the control and primed seeds under observation showed marked differences in the timing of initiation and completion of germination.
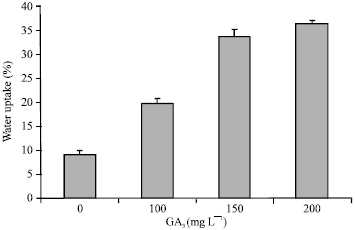 | |
| Fig. 1: | Water uptake of control and primed seed soaked for 2 h in 20 mL of distilled water |
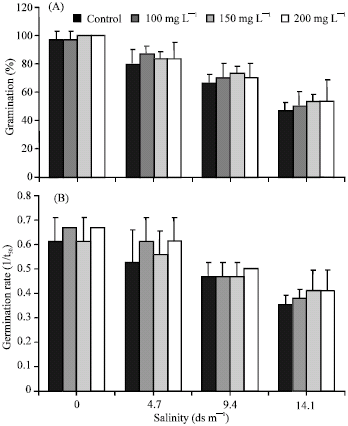 | |
| Fig. 2: | Germination (A) and germination rate (B) of sugar beet seeds primed with 0.0, 100, 150 and 200 mg L-1 GA3 under various salt concentrations |
Germination started within 36 h and was complete on the 8th day. Seed germination delayed as the level of salinity increased. However, priming treatments shortened the time to seed germination (Fig. 2B). Figure 2B also showed seeds primed with 150 and 200 mg L-1 GA3 took less time to complete germination at high salt concentration as compared to other treatments.
Further studies were carried out to observe the effect of salt stress on early seedling growth of germinating seeds of different priming treatments. The results indicated that an increased salt concentration caused delayed emergence of root and shoot compared to primed seeds. A continuous increase in length of root and shoot was observed in frequent hours of germination in the control as well as primed seeds under salt treatments. The average length of root and shoot (Fig. 3) shows that all treatments showed a strong inhibition with the increasing level of salt solution. There was considerable reduction in the size of root and shoot at highest level of salinity (14.1 dS m-1 NaCl). However, primed seeds showed less reduction in the size of root and shoot as compared to control (Fig. 3). Maximum root length was measured at 100 and 200 mg L-1 GA3 under high salt concentrations while maximum shoot length was measured at 100 and 150 mg L-1 GA3. Plants raised from seeds treated with 100 mg L-1 GA3 had the maximum shoot length under both the salt and control treatments (Fig. 3).
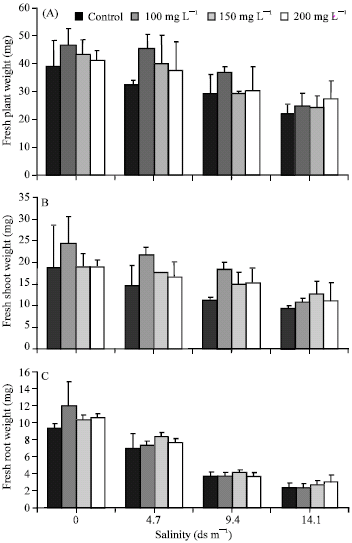
Increase in salt concentration of the growth medium caused a significant reduction in mean fresh weights of plants, roots and shoots. Higher fresh weights were recorded from primed seeds as compared to control (Fig. 4).
 | |
| Fig. 3: | Root length (A) and shoot length (B) of sugar beet plants raised from seeds primed with 0.0, 100, 150 and 200 mg L-1 GA3 under various salt concentrations |
| |
| Fig. 4: | Fresh weights of plants (A), shoots (B) and roots (C) of sugar beet plants raised from seeds primed with 0.0, 100, 150 and 200 mg L-1 GA3 under various salt concentrations |
Plants raised from seeds treated with 100 mg L-1 GA3 had the greater fresh weights of plants and shoots under both the salt and control treatments except 14.1 dS m-1 NaCl. Whereas greater fresh weights of roots were recorded at 200 mg L-1 GA3 while maximum fresh weights of shoots were observed at 150 mg L-1 GA3 under highest salt concentrations (Fig. 4).
DISCUSSION
It is evident that plant growth regulators have been found to play a central role in the integration of the responses expressed by plants under stress conditions (Amzallag et al., 1990). Gibberellic acid has been reported to increase percentage germination and seedling growth (Kaur et al., 1998). In this study, primed seeds show enhanced performance under stress conditions. Seed priming shortened germination time, but stress conditions delayed it considerably (Fig. 2B). Furthermore, priming resulted in increase of normal germination in all primed seeds. This could be explained by more rapid water uptake in primed seeds because germination in primed seeds started after 24 h. It supports that priming caused more rapid water uptake than control (Fig. 1). These results agree with Murillo-Amador et al. (2002) in cowpea, Demir and Van De Venter (1999) in watermelon, they suggested that salinity may influence germination by decreasing the water uptake. The pre-sowing treatments cause initiation of the early metabolic processes and the re-drying of seeds arrest, but do not reverse, the initial stages of germination so that on the availability of suitable conditions, the time taken to germinate is reduced (Bewley and Black, 1982). Priming-induced activation of metabolic events (Hanson, 1973) has been reported in seeds of various plant species. Akinola et al. (2000) reported that higher duration of exposure to seed treatment resulted in higher cumulative germination in wild sunflower and Caseiro et al. (2004) found that hydropriming was the most effective method for improving seed germination of onion, especially when the seeds were hydrated for 96 h compared to 48 h. The results are in line with the findings of Thornton and Powell (1992) in Brassica and Srinivasan et al. (1999) in mustard.
The root and shoot length are the most important parameters for salt stress because roots are in direct contact with soil and absorb water from soil and shoot supply it to the rest of the plant. For this reason, root and shoot length provides an important clue to the response of plants to salt stress (Jamil and Rha, 2004). It was observed that priming improved root and shoot growth as compared to control (Fig. 3). Higher fresh weights of plants, shoots and roots were also recorded from primed seeds as compared to control (Fig. 4). Major effect of seed treatment on seedling growth observed was due to faster emergence, giving seedlings a longer time to develop (Parera and Cantliffe, 1994). Muhyaddin and Wiebe (1989) suggested that the enzymes are activated with an accompanying mobilization of reserve materials ending in transport of the reserve materials in the embryo by osmotic conditioning and thus stronger seedlings are obtained as a result of embryo growth. There are numerous studies in the literature which exhibit the considerable effectiveness of hydro-priming on germination and later growth in different plant species under both saline and non-saline conditions, e.g., wheat (Idris and Aslam, 1975), Acacia tortilis and Acacia coriacea (Rehman et al., 1998) and only under normal non-saline conditions e.g., maize (Zea mays), rice (Oryza sativa) and chickpea (Cicer arietinum) (Harris et al., 1999).
ACKNOWLEDGMENT
The present research was conducted by the research fund of Rural Development Administration (RDA), Korea in 2006.
REFERENCES
- Akinola, J.O., A. Larbin, G.O. Farinu and A.A. Odunsi, 2000. Seed treatment methods and duration effects on germination of wild sunflower. Exp. Agric., 36: 63-69.
CrossRef - Amzallag, G.N., H.R. Lerner and A. Poljakoff-Mayber, 1990. Exogenous ABA as a modulator of the response of Sorghum to high salinity. J. Exp. Bot., 541: 1529-1534.
CrossRefDirect Link - Caseiro, R., M.A. Bennett and J. Marcos-Filho, 2004. Comparison of three priming techniques for onion seed lots differing in initial seed quality. Seed Sci. Technol., 32: 365-375.
Direct Link - Demir, I. and H.A. Van-De-Venter, 1999. The effect of priming treatments on the performance of watermelon (Citrillus lanatus (Thunb.) Matsum. and Nakai) seeds under temperature and osmotic stress. Seed Sci. Technol., 27: 871-875.
Direct Link - Hanson, A.D., 1973. The effects of imbibition drying treatments on wheat seeds. New Phytol., 72: 1063-1073.
CrossRefDirect Link - Harris, D., A. Joshi, P.A. Khan, P. Gothkar and P.S. Sodhi, 1999. On-farm seed priming in semi-arid agriculture: Development and evaluation in maize, rice and chickpea in India, using participatory methods. Exp. Agric., 35: 15-29.
Direct Link - Idris, M. and M. Aslam, 1975. The effect of soaking and drying seeds before planting on the germination and growth of Triticum vulgare under normal and saline conditions. Can. J. Bot., 53: 1328-1332.
CrossRefDirect Link - Jamil, M. and E.S. Rha, 2004. The effect of salinity (NaCl) on the germination and seedling of suger beet (Beta vulgaris L.) and cabbage (Brassica oleracea capitata L.). Korean J. Plant Res., 7: 226-232.
Direct Link - Kaur, S., A.K. Gupta and N. Kaur, 1998. Gibberellic acid and kinetin partially reverse the effect of water stress on germination and seedling growth in chickpea. Plant Growth Regul., 25: 29-33.
CrossRefDirect Link - Khajeh-Hosseini, M., A.A. Powell and I.J. Bingham, 2003. The interaction between salinity stress and seed vigour during germination of soyabean seeds. Seed Sci. Technol., 31: 715-725.
CrossRefDirect Link - Murillo-Amador, B., R. Lopez-Aguilar, C. Kaya, J. Larrinaga-Mayoral and A. Flores-Hernandez, 2002. Comparative effects of NaCl and polyethylene glycol on germination, emergence and seedling growth of cowpea. J. Agron. Crop Sci., 188: 235-247.
CrossRefDirect Link - Parera, C.A. and D.J. Cantliffe, 1994. Presowing Seed Priming. In: Horticultural Reviews, Volume 16, Janick, J. (Ed.). John Wiley and Sons, Oxford, UK., pp: 109-141.
CrossRef - Rehman, S., P.J.C. Harris and W.F. Bourne, 1998. Effect of pre-sowing treatment with calcium salts, potassium salts or water on germination and salt tolerance of Acacia seeds. J. Plant Nutr., 21: 277-285.
Direct Link - Sadeghian, S.Y. and N. Yavari, 2004. Effect of water-deficit stress on germination and early seedling growth in sugar beet. J. Agron. Crop Sci., 190: 138-144.
CrossRefDirect Link - Santa-Cruz, A., M.M. Martinez-Rodriguez, F. Perez-Alfocea, R. Romero-Aranda and M.C. Bolarin, 2002. The rootstock effect on the tomato salinity response depends on the shoot genotype. Plant Sci., 162: 825-831.
CrossRefDirect Link - Singh, B.G., 1995. Effect of hydration-dehydration seed treatments on vigour and yield of sunflower. Indian J. Plant Physiol., 38: 66-68.
Direct Link - Sivritepe, H.O. and A.M. Dourado, 1995. The effect of priming treatments on the viability and accumulation of chromosomal damage in aged pea seeds. Ann. Bot., 75: 165-171.
CrossRefDirect Link - Srinivasan, K., S. Saxena and B.B. Singh, 1999. Osmo-and hydropriming of mustard seeds to improve vigour and some biochemical activities. Seed Sci. Technol., 27: 785-789.
Direct Link - Jamil, M., E.S. Rha and M.S. Han, 2006. Salinity effect on ion content and relationship between ion leakage, growth and salinity in different vegetable seeds. Seed Technol. J., 28: 47-57.
Direct Link