
Muhammad Zaffar Iqbal
Not Available
Muhammad Aslam Nadeem
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 4 | Page No.: 395-399







ABSTRACT
Genic effects for yield of seed cotton and number of sympodial branches per plant were estimated from two Upland cotton crosses through generation mean analysis from six populations (P1, P2, F1, F2, BC1 and BC2). The results showed that 5 crosses over mid and 4 crosses over better parent showed significant heterosis for number of sympodial branches per plant, whereas only 4 crosses exhibited inbreeding depression for this character. The generation mean analysis advocates the presence of additive gene action in crosses i.e., S-12 x S-14, S-12 x Albacala (69)11 , LRA-5166 x S-12 and LRA-5166 x S-14 for number of sympodial branches per plant. All the crosses exhibited heterosis in desired direction over mid and better parents except Albacala (69)11 x S-12 for yield of seed cotton plant -1 Significant marked inbreeding depression from F1 to F2 generation were observed in all the crosses except S-14 x LRA5166 for yield of seed cotton per plant. Maximum degree of dominance was associated with S-14 x S-12. It reflected the presence of maximum number of dominant genes, which needs to be the cause of significant heterosis in this combination. Most recessive alleles for seed cotton yield accumulated in Albacala (69) 11 x S-12, which expressed least positive degree of dominance. The scaling test revealed involvement of epistasis in all the crosses, except S-14 x LRA-5166 for yield of seed cotton per plant. The rest of all the crosses were predominately under non additive genetic control except S-14 x LRA5166 for yield of seed cotton plant -1, hence delayed selection will be fruitful in these crosses. However the presence of duplicate epistasis in all the crosses would limit the pace of variability and thus slow down the pace of progress through selection. Therefore, heterosis breeding would be advantageous for the improvement of yield of seed cotton per plant.
PDF Abstract XML References Citation
How to cite this article
Muhammad Zaffar Iqbal and Muhammad Aslam Nadeem, 2003. Generation Mean Analysis for Seed Cotton Yield and Number of Sympodial Branches per Plant in Cotton (Gossypium hirsutum L.). Asian Journal of Plant Sciences, 2: 395-399.
DOI: 10.3923/ajps.2003.395.399
URL: https://scialert.net/abstract/?doi=ajps.2003.395.399
DOI: 10.3923/ajps.2003.395.399
URL: https://scialert.net/abstract/?doi=ajps.2003.395.399
INTRODUCTION
The different types of gene actions are important in different crosses. The breeding strategy should therefore, be based on the gene action involved in that particular cross to get a desirable genotype.
Some researchers from abroad like Saudhu and Nittal (1988) studied two G. arborieum crosses and reported absence of non allelic interaction in six parameter model while three parameter model predicted, the presence of epistasis for yield of seed cotton per plant. One cross had significant additive, additive x additive and dominance x dominance while the other cross had only significant additive effects. Ketageri et al. (1992) showed significant positive heterosis over mid parent and high magnitude of inbreeding depression for yield of seed cotton per plant. Iqbal (1994) performed and Hayman (1954) reported the genetic analysis and observed epistasis in F1 and F2 generation of 9 x 9 diallel crosses for yield of seed cotton per plant. He also advocated the presence of both additive and non additive genetic variation for yield of seed cotton per plant. Azhar et al. (1994) through Hayman analysis advocated involvement of epistasis in the inheritance of number of sympodial branches per plant. Khan et al. (1996) reported that high yielding parents did not produce high yielding hybrids. Yingxin and Xiangming (1998) and Abdallah et al. (1999) reported the presence of both additive and non-additive genetic effect for yield of seed cotton. Using Griffing (1956) technique Subhan et al. (2001) observed significant differences among hybrids and their parents for yield of seed cotton per plant.
In Pakistan on cotton plant no body had carried out generation mean analysis probably due to creation of six generations and plantation of these generations in one year to overcome the problem of genotypic environment interaction. Keeping this issue in mind this study was undertaken to know the breeding behavior, nature and magnitude of gene effects controlling seed cotton yield and number of sympodial branches in Upland cotton.
MATERIALS AND METHODS
Four diverse parental genotypes of Upland cotton viz., S-12, S-14, Albacala (69)11 and LRA5166 were selected and selfed for two generations before being used in a diallel crossing. These four parents were crossed in diallel fashion during 1998-99. In 1999-2000 these parents were again crossed in diallel technique and F1 was used for the creation of F2 and back crossing with female (Bc1) and male (Bc2) parents. The four parents, their F1, F2, BC1 and BC2 generation were grown in a randomized complete block design (RCBD) with three replicated during 2000-01 at Cotton Research Sub Station, Khanewal with intra and inter row distance of 30 and 75 cm, respectively. The F1, P1, P2, BC1 and BC2 were presented by three rows of 4.3 meter each per replication while F2 were planted on 4.3 x 6 m plot in each replication. Yield of seed cotton per plant and number of sympodial branches were recorded for each and every plant in three replications. The data were subjected to analysis of variance given by Steel and Torrie (1980). Heterosis and heterobeltosis were computed in terms of percentage increase (+) or decrease (-) of F1 over mid and better parent respectively. The potence ratio was calculated, method proposed by Griffing (1950). The inbreeding depression was computed in terms of percentage decrease at F2 over F1. The environmental, additive and dominance genetic variance (three parameter model) were computed as described by Simmonds (1979). The scaling tests were performed following Mather and Jinks (1989) and Hayman and Mather (1955) in order to determine the presence or absence of non allelic gene interaction. Six parameters model suggested by Hayman (1958) were used for estimation of various genetic components, assuming the absence of linkage, multiple alleles, lethal genes and full viability of gametes and zygotes.
Results and Discussion
Yield of seed cotton per plant: The genotypes differed significantly in their genetic constitution (Table 1). This genetic variability paves the way for further analysis.
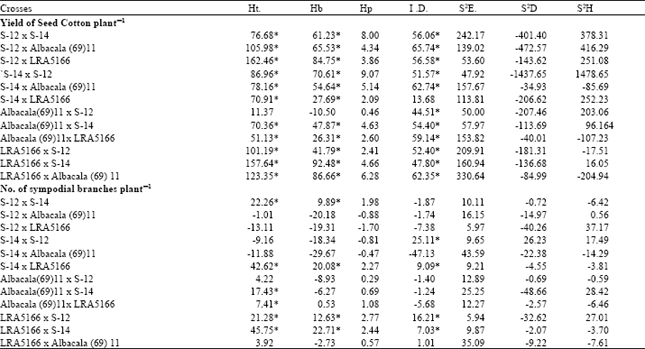
The estimation of heterosis (Table 2) based on F1 generation mean over mid parental values were significant is desired direction for all the crosses except Albacala (69) 11 x S-12. These results are in accordance with the findings of Subhan et al. (2001). The maximum heterosis 162.46% was noted in cross combination S-12 x LRA5166. All the crosses exhibited significant heterosis over better parents except Albacala (69) 11 x S-12. The heterobeltosis ranged from –10.5% (LRA5166 x S-14) to 92.48% (Albacala (69) 11 x S-12). Although the high yielding parents produced significant heterosis but it was noted that in both heterosis and heterobeltosis higher yielded crosses combination had one lower yielder parent. These findings are in agreement with Khan et al. (1996) who observed that high yielding parent did not produce high yielding hybrid. The heterosis in reciprocal crosses were not in same direction, hence the heterosis was greatly effected by the direction in which way the cross was made.
| Table 1: | Mean square of replication, genotypes and error for yield of seed cotton and No. of sympodial branches |
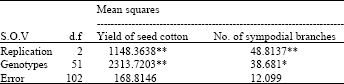 | |
| * = Significant at 5% probability level. | |
| ** = Significant at 1% probability level. | |
Significant marked inbreeding depression from F1 to F2 generation were observed in all the crosses except S-14 x LRA-5166, hence its exploitation in selecting good segregate could be made. These results are in accordance with the findings of Ketageri el al. (1992). The high dominance estimate in the form of potence ratio indicated preponderance of dominance genetic variations in 11 cross combinations except Albacala(69)11 x S-12. Iqbal (1994), Yingxin and Xiangming, (1998) and Abdallah et al. (1999) observed both additive and non additive effect for this character. This contradiction may be due to different methods used for analysis. The generation means analysis is more reliable than simple additive dominance model. Maximum degree of dominance was associated with S-14 x S-12 and its reciprocal. It reflects the presence of maximum number of dominant alleles, which needs to be the cause of significant heterosis in this combination. Most recessive alleles for seed cotton yield accumulated in Albacala (69)11 x S-12 which expressed least positive degree of dominance. The three parameter model indicated the importance of environment for character expression in all the crosses, whereas dominance genetic variance (S2H) was important for all the crosses except S-14 X Albacala (69)11, Albacala (69)11 x LRA5166 its reciprocal and LRA5166 x S-12.
The results of scaling tests revealed significant differences for one or the other tests in all the crosses except S-14 x LRA5166 (Table 3). These significant tests indicated inadequacy of simple additive – dominance model, hence six parameters model was needed to explain the genetic variation. The scaling test also suggested the major role of epistasis for controlling yield of seed cotton per plant.
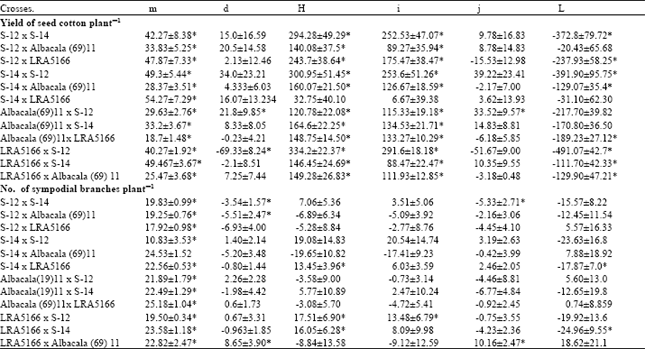
The perusal of data indicated significant dominance component (h) (Table 4), which revealed that yield of seed cotton per plant was predominantly under non-additive genetic control in all the crosses except S-14 x LRA5166. The high magnitude of dominance effect suggested that selection of high yielding genotypes would be postponed till later generation when the dominance effect would have diminished except S-14 x LRA5166, Albacala (69)11 x S-12 and LRA5166 x S-12.
In cross combination Albacala (69)11 x S-12 and LRA5166 x S-12 the additive genetic effect (d) was equally important as non-additive, therefore, for efficient utilization of fixable and unfixable components of genetic variation, reciprocal recurrent breeding method was suggested for these crosses.
| Table 2: | Heterosis, Heterobetoiosis, Potence ratio, Inbreeding depression and Genetic variance, Environmental, Additive and Dominance) for Yield of seed cotton and No. of sympodial branches plant-1 in Gossypium hirsutum crosses |
 | |
| Table 3: | Scaling test for Yield of Seed cotton and Number of Sympodial branches in cotton |
 | |
| Table 4: | Estimates of components of generation mean analysis for yield of seed cotton and Number of sympodial branches per plant in Gossypium hirsutum L. Crosses alongwith their standard errors |
 | |
As this breeding procedure will concentrate additive effect, but will not allow to dissipate non-additive effect.
The results also revealed that additive x additive interaction (I) contributes significantly in all the crosses except S-14 x LRA5166, these findings were corroborated with scaling test which also indicated the absence of interaction in this cross. The additive x dominance type of gene interaction (j) was significant in two crosses i.e., Albacala (69)11 x S-12 and LRA5166 x S-12. The dominance x dominance type of gene interaction (I) contributed significantly in all the crosses except S-12 x Albacala (69)11 and S-14 x LRA5166. These findings are in accordance with Saudhu and Nittal (1988). The opposite sign of (h) and (I) in all the crosses suggested duplicate type of epistasis, which would limit the range of variability and thus slow down the pace of progress through selection, therefore, hetorosis breeding would be advantageous.
Number of sympodial branches plant-1: The analysis of variance for experiment (Table 1) showed significant genotypic differences, which implies that there was substantial amount of genetic variability among the genotypes for further study.
Table 2 indicated that heterosis varied from –13.105 percent (S-12 x LRA5166) to 45.747 percent (LRA5166 x S-14) and heterobeltosis from –29.662 % (S-14 x Albacala (69)11) to 22.71 % (LRA5166 x S-14) (Table 2). The significant inbreeding depression was noted in four crosses and three of them had positive significant heterosis and heterobeltosis. The potence ratio was higher for only three crosses. The results also revealed that cross combination S-14 x S-12 had higher both additive and dominance genetic variance whereas the only dominance genetic variance was higher in S-12 x LRA5166, Albacala (69)11 x S-14 and LRA5166 x S-12 crosses (Table 2). The higher environmental variance for test of the crosses indicted the importance of environment in character expression of number of sympodial branches plant-1.
The results of scaling test divulged the absence of non-allelic interaction in most of the crosses except S-12 x S-14, S-14 x LRA5166, LRA5166 x S-12 and LRA5166 x S-14 (Table 3). These findings partially agreed with the results of Azhar et al. (1994). This non- interacted trend in majority of crosses revealed adequacy of additive– dominance model to account for the analysis of data.
Additive genetic effect (d) were significant in the crosses S-12 x S-14, S-12 x Albacala (69)11 and LRA5166 x Albacala (69)11. The dominant genetic effects (h) were significant for S-14 x LRA5166, LRA5166 x S-12 and LRA5166 x S-14 (Table 4). Epistasis interaction (I) was presented for only one cross LRA5166 x S-12 while additive x dominance gene interaction (j) were noted for S-12 x S-14 and LRA5166 x Albacala (69)11. Dominance type of gene interaction was found in crosses S-14 x LRA5166 and its reciprocal.
REFERENCES
- Azhar, F.M., N. Khan and S.K. Ajmal, 1994. Genetic basis of variation in upland cotton. J. Agric. Res., 32: 9-16.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Hayman, B.I., 1960. The separation of epistatic from additive and dominance variation in generation means. II. Genetica, 31: 133-146.
CrossRefDirect Link