
R. Karthika
Not Available
B. Subba Lakshmi
Not Available
Asian Journal of Plant Sciences
Year: 2006 | Volume: 5 | Issue: 4 | Page No.: 721-724







ABSTRACT
A study was undertaken on induced mutagenesis with CO1 and CO2 varieties of soybean. The mutagens studied were gamma rays (20, 30, 40, 50 and 60 kR) and EMS (5, 10, 15, 20 and 25 mM). In an assessment to fix the LD50 value, the varieties responded differently. A subtle difference was observed between the varieties CO1 and CO2 in the degree of tolerance to the mutagens, viz., CO1, (62 kR gamma rays, 26.4 mM EMS) and CO2 (58.3 kR gamma rays and 25.7 mM EMS). It is concluded that CO1 tolerate higher doses of gamma rays and EMS. In general, the treated population had manifested a reduced expression than the untreated population in all the biometrical traits. Reduction in phenotypic expression is linear with the dose or concentration of the mutagens in M1 generation. The mutagen gamma ray manifested higher percentage of reduction than EMS. In M2, EMS induced higher proportion of chlorophyll mutants than gamma rays. Among viable mutants, sterile mutant occurred more predominantly followed by stunted growth in both the varieties due to the effect of mutation. Induced mutations delivered fairly good amount of genotypic coefficient of variation, the heritability, GA as percent of mean with respect to number of seeds per plant, seed yield per plant and hundred seed weight for both the mutagen in both the varieties, indicating the scope for improving the soybean yield by selection. The mutants with four seeded pods, bold seeded types and increased number of pods per plant were isolated from this experiment.
PDF Abstract XML References Citation
How to cite this article
R. Karthika and B. Subba Lakshmi, 2006. Effect of Gamma Rays and EMS on Two Varieties of Soybean. Asian Journal of Plant Sciences, 5: 721-724.
DOI: 10.3923/ajps.2006.721.724
URL: https://scialert.net/abstract/?doi=ajps.2006.721.724
DOI: 10.3923/ajps.2006.721.724
URL: https://scialert.net/abstract/?doi=ajps.2006.721.724
INTRODUCTION
Soybean (Glycine max (L.) Merrill.) tops the world production of oilseeds. The productivity of soybean in India is much low in comparison with the world average. The attributes identified for such low productivity are limited genetic diversity, narrow genetic base of Indian soybean varieties, short growing period available in Indian latitude, stagnant genetic potential for yield (Tiwari, 2003). The narrowing down of the genetic base is due to the repeated use of few parents for breeding programme (Satyavathi et al., 2003). At this context, the widening of the genetic base is a major concern and challenge put forward to the Indian soybean breeders.
In soybean, creation of variation through hybridization is a tedious process, due to small, fragile flowers that, makes it difficult to carry out the process of emasculation injuring the parts of the flower and are prone to heavy flower shedding (over 75%) even under favorable conditions (Johnson and Bornard, 1976). These coupled with complete self- fertility impose limitation on the success of hybridization programme. Hence the classical breeding methods have got limited application in soybean and as such, mutation breeding appears to play an important role in the improvement of this important pulse-cum oil seed crop.
MATERIALS AND METHODS
Two varieties CO1, CO2 of soybean (Glycine max (L.) Merill) formed the materials for the present investigation. The investigation envisaged studying the differential sensitivity of the soybean varieties by subjecting them to different doses of the two mutagens-Gamma rays (Physical mutagen) and ethyl methane sulphonate (Chemical mutagen). Gamma irradiation was done using the cobolt 60 sources in the gamma chamber, installed at the Sugarcane Breeding Institute, Coimbatore. The chemical mutagen, ethyl methane sulphonate (CH3SO2OC2H5) with a molecular weight 124.16, from the sigma chemical company, USA was used for treating the seeds.
For the assessment of LD50 dose two hundred seeds of uniform size were used for (Gamma rays - 0, 20, 40, 60, 80, 100 kR and EMS - 0, 5, 10, 15, 20, 25 and 30 mM ) each treatment. In respect of EMS treatment, the seeds were presoaked in double distilled water for 6 h. Appropriate quantities of EMS were dissolved in double distilled water to have the concentrations envisaged in the programme. The treatment was performed at room temperature of 22±2°C at early morning hours with intermittent shaking during the treatment period of 6 h. After the chemical treatment, the seeds were washed thoroughly with running tap water for half an hr to remove the residues of the chemical, if any and the excess moisture in seed coat was removed by using folds of blotting paper.
The treated seeds with control were sown in germination trays at Germination room, Seed science and technology department, Tamil Nadu Agricultural University, Coimbatore. The seeds were sown in three replications at the rate of 65 seeds per replication in 30x30 spacing. The following observations such as germination and survival percentage on 30th day were recorded. For working out the lethal dose (50%) probit analysis was done using the germination and survival percentage (Sharma, 1998).
One eighty seeds in each of the varieties CO1 and CO2 were treated with gamma rays at 20, 30, 40, 50 and 60 kR and EMS at 5, 10, 15, 20 and 25 mM concentrations for each treatment. The treated seeds with control were sown in Millet Breeding Station, TNAU, Coimbatore, with three replications adopting a spacing of 30x30 cm. In each replication three rows were sown with 20 seeds per row for each treatment.
The materials used in M1 are forwarded to M2 generation as progeny to row method (Plants yielded > 40 seeds). The experiment was laid out in an unreplicated trial at spacing of 30x30 cm.
RESULTS
LD50 is of great importance to know the sensitivity of different genotypes to the critical dose of mutagens causing 50% mortality. Probit analysis was done based on the survival of seedlings in both the varieties for both the mutagens to determine the Lethal Dose (LD50) dose. According to the results the LD50, for CO1 and CO2 were worked as 62 and 58.3 kR (Gamma rays) and the LD 50 for CO1 and CO2 were 26.40 and 25.70 mM (EMS), respectively. A slight variation in LD50 between different varieties of same species is a common phenomenon in mutation breeding as the resistance offered by the biological material depends on size, maturity, hardness and moisture content etc., at the time of treatment. Such variation was noticed in black gram by Vanniarajan and Vijendra Das (1996).
In this investigation, the germination percentage and plant survival were found to be inhibited by irradiation and EMS for both the varieties. The percentage of seed germination and survival progressively decreased with increasing dose or concentration of mutagens. Reduction in survival was also due to reduced primary leaf growth and resultant death of seedlings. Besides the highly injured plant (50 kR in CO1), the other abnormalities noticed in the M1 population were reduced primary leaf (60 kR gamma rays and 25 mM EMS in CO2), malformed cotyledonary leaves (15 mM in CO1). Such decrease in germination, survival and seedling growth due to mutagen observed in the present study are in conformity with the earlier reports of Balakrishnan (1991) in soybean.
In M2 generation progressive enhancement in the frequency of chlorophyll mutations with increasing doses of both the mutagen was observed. EMS induced higher proportion of chlorophyll mutants than gamma rays. Gamma rays and EMS produced four types of chlorophyll mutants viz., albina, chlorina, Xantha and viridis. In EMS treated population, xantha occurred more frequently followed by chlorina, albina nd viridis. In gamma treated population albina occurred more frequently in both the varieties. This was in agreement with the results obtained by Singh et al. (2001) in rice. Xing et al. (1997) opined that chloroplasts in albino leaves were normal but the inner structure was simpler and lacked a rich and highly organized internal thylakoid system. Only stroma and DNA- like fibrils were observed. The development of chloroplast is thus obviously delayed leading to chlorophyll deficient mutants.
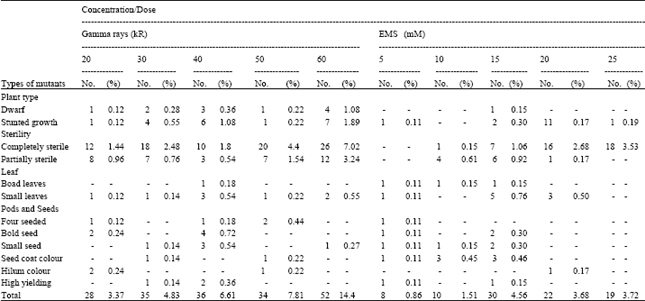
The spectrum of viable mutations observed in the study consisted of dwarf, stunted growth, leaf aberrant, sterile mutants, variation for seed characters and high yielding types. More number of viable mutants was recorded in gamma treated population of CO1 and EMS treated population of CO2 (Table 1 and 2). The frequency of viable, non-viable and total mutation on M2 plant basis showed inconsistent relationship with mutagen. Among the number of viable mutations for different traits, sterile mutant occurred more predominantly followed by mutants with stunted growth in both the varieties. Mutation induced male sterility in soybean was due to degeneration of generative cell at pollen meiosis II which resulted in low number of viable pollens (Bione et al., 2002). Partial sterile mutants were common in all the treatments in both the varieties. Groose and Palmer (1987) reported that partial sterility in soybean was characterised by a reduced number of seeds per pod, which appeared to be the result of ovule or very early embryo abortion.
High yielding mutants were observed in both the varieties. Plants showing increased vigour and yield of pods coupled with highest 100 seed weight have been found among them. Such increase in vigour and yield have been observed by Pushpendra and Ram (1990) in gamma-irradiated population of soybean.
| Table 1: | Spectrum of viable mutants in CO1 population |
 | |
| Table 2: | Spectrum of viable mutants in CO2 population |
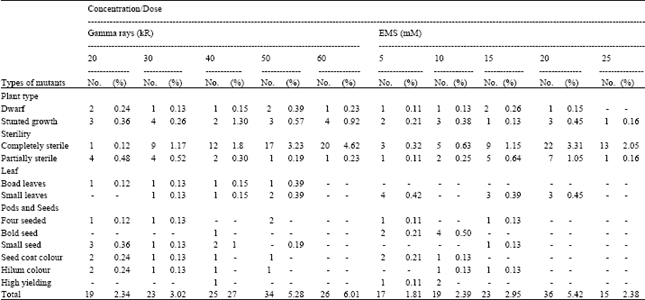 | |
Black discolouration of hilum was noticed in both the varieties. Takahashi and Abe (1994) reported yellow hilum in soybean. Cober et al. (1998) assessed that the colour of the hila ranges from yellow to brown depending upon the genetic background of the line and environmental conditions. Productive characters like number of seeds per plant, seed yield per plant, hundred seed weight exhibited low variation between phenotypic and genotypic co efficient of variation and that too predominant at 15 mM in CO1 and 40 kR gamma rays in CO2. So these two varieties at specified dosages could be emphasized in selection programme.
REFERENCES
- Cober, E.R., G.R. Ablett, R.I. Buzzell, B.M. Luzzi, V. Poysa, A.S. Sahota and H.D. Voldeng, 1998. Imperfect yellow hilum colour in soybean is conditioned by II rr TT. Crop Sci., 38: 940-941.
Direct Link - Groose, R.W. and R.G. Palmer, 1987. Partially sterile mutants in a genetically unstable line of soybean. Soybean Genet. Newslett., 14: 169-171.
Direct Link - Pushpendra and H.H. Ram, 1990. Dry matter yield as an effective selection criterian in soybean. Trop. Agric., 67: 57-60.
Direct Link - Singh, S., J. Singh and R.K. Singh, 2001. Gamma ray, EMS and sodium azide induced effectiveness and efficiency of chlorophyll mutations in Basmati rice (Oryza sativa L.). Crop Res., 22: 113-120.
Direct Link - Takahashi, R. and J. Abe, 1994. Genetic and linkage analysis of low temperature induced browning in soybean seed coats. J. Hered., 85: 447-450.
Direct Link - Tiwari, S.P., 2003. Improvement of yield and yield potential in soybean: An analysis and synthesis. J. Oilseeds Res., 20: 1-8.
Direct Link - Vanniarajan, C. and L.D. Vijendra-Das, 1996. Induced variation for seed protein content in Blackgram. Madras Agric. J., 84: 114-115.
Direct Link