
Emtithal M. Abd-El-Samie
Department of Entomology, Faculty of Science, Cairo University, Egypt
Hoda B. Faheem
Department of Cell Biology, National Research Center, Egypt
Mary T. Ibrahim
Department of Cell Biology, National Research Center, Egypt
Adel Ramzy
Department of Entomology, Faculty of Science, Ain Shams University, Egypt
Mohammed S. Salama
Department of Entomology, Faculty of Science, Ain Shams University, Egypt
Biotechnology
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 364-372







ABSTRACT
Drosophila melanogaster is a widely used model organism for genetic dissection of developmental processes. It is an ideal multicellular organism for the rapid toxicological screening of substances for mutagenicity. Boule gene was shown to be a key regulator of meiosis in Drosophila. Boule-deficient fruit flies are infertile and display meiotic arrest in the male germ cells. Investigation of the impact of some nitrogenous fertilizers on the expression of Drosophila boule gene had been performed. Newly hatched larvae were treated with the LC25 and LC50 of the three tested nitrogenous fertilizers Urea, Ammonium nitrate and Calcium nitrate. The study includes the isolation and molecular characterization of the Drosophila boule gene homolog to the human DAZ gene. Total RNA have been isolated from untreated and treated D. melanogaster adult male flies. Fragments of the boule gene were recovered with RT-PCR for sequencing. Treated and untreated Drosophila melanogaster boule gene express the same pattern of transcripts. The sequence of the boule fragments of the treated adult males with the LC25 and LC50 of the three tested nitrogenous fertilizers showed different types of mutations such as substitution, deletion and insertion mutations. These results indicate that the three nitrogenous fertilizers used posses mutagenic potentialities. The mutagenic activity is due to their Nitrogen (N) content. Another possible interpretation for the mutagenicity of the tested compounds is the ability of nitrogenous fertilizers to methylate DNA resulting in different types of genetic alterations. Analysis of these mutations should provide insight into the genetic networks that control male fertility in Drosophila and other organisms, including humans.
PDF Abstract XML References Citation
How to cite this article
Emtithal M. Abd-El-Samie, Hoda B. Faheem, Mary T. Ibrahim, Adel Ramzy and Mohammed S. Salama, 2007. Molecular Analysis for Nitrogenous Fertilizers Effect on Drosophila melanogaster Boule Gene. Biotechnology, 6: 364-372.
DOI: 10.3923/biotech.2007.364.372
URL: https://scialert.net/abstract/?doi=biotech.2007.364.372
DOI: 10.3923/biotech.2007.364.372
URL: https://scialert.net/abstract/?doi=biotech.2007.364.372
INTRODUCTION
All living systems need nitrogen for the production of complex organic molecules, such as proteins, nucleic acids, vitamins, hormones and enzymes. Due to the intense use of synthetic nitrogenous fertilizers and livestock manure in modern day agriculture, food (particularly vegetables) and drinking water may contain higher concentrations of nitrate than in the past. Nitrate may also be synthesized endogenously from nitric oxide, which reacts to form nitrite. Nitrate itself is generally regarded nontoxic. Toxicity is usually the result of the conversion of nitrate into the more toxic nitrite. There are two major toxicological concerns regarding nitrite. First, nitrite may induce methaemoglobinaemia, which can result in tissue hypoxia and possibly death. Secondly, nitrite may interact with secondary or N-alkyl amides to form N-nitroso carcinogens (Mensinga et al., 2003). Owing to the risk arising from the continuous use of increased quantities of nitroso compounds, such as nitrogenous fertilizers, the human environmental hazard is the most important problem for public health. United Nation (1976).
Recent studies showed that nitrogenous fertilizers have various hazards on the genetic material. These hazards can appear as malignancy in the actual living populations or may represent genetic load to future generation (Xu et al., 2003).
Sodium nitrite is one of the most important pollutant agents used as a food additive. Food and Drug Administration (FDA) studies showed its toxicity and carcinogenesis (International Toxicol Program Ser., 2001). Sodium nitrite causes various harmful effects in different animals: In rats and mice, it causes brown discoloration in the eyes and cyanosis of the mouth, tongue, ears and feet of males. Reticulocyte counts increased in males and females. Methaemoglobin concentrations were elevated in almost all exposed groups. Sperm motility decreased significantly (Miasoedova and Nazarov, 2004). The genotoxicity of endogenously formed N-nitrosoamines from secondary amines and sodium nitrite (NaNO2) was evaluated in multiple organs of mice (Ohsawa et al., 2003).
In chickens, it causes pharyngo-esophageal carcinoma, gastric adenocarcinoma and liver carcinoma (Xu et al., 2003).
Urea is a protein unfolding agent that can accumulate locally in high concentrations in tissues of many organisms. David et al. (1999) used Drosophila melanogaster to test the hypothesis that urea loading would promote the formation of isoaspartate (beta-carboxyl-linked aspartate ) a common form of protein damage that occurs most readily in unstructured polypeptides and flexible regions of folded proteins. Also, N-ethyl-N-Nitrosourea (ENU) was mutagenic in all stages of oogenesis in female of Drosophila melanogaster, although there were differences among the stages. Studies with female germ cells could be an alternative to the use of premeiotic male germ cells. Moreover, the molecular spectrum induced with ENU was studied in pre-meiotic repair active male germ cells of Drosophila melanogaster (Alvarez et al., 2002).
In Drosophila spermatogenesis, meiotic cell cycle progression and cellular differentiation are linked by the function of the meiotic arrest genes. The meiotic arrest genes control differentiation by regulating the transcriptional activation of many specific genes. The meiotic arrest genes have been subdivided into: Always early (aly) and cannonball (can) classes, based on the mechanism by which they control cell cycle progression. A number of meiotic regulators such as cyclin B, boule and twine, as well as many genes required for spermatid differentiation, are under the control of meiotic arrest genes (White-cooper et al., 2000; Perezgasga et al., 2004).
Drosophila genes: doublesex, boule and diaphanous share extensive sequence with human genes that cause male sex reversal, lack of sperm and premature ovarian failure, respectively. These corespondences could lead to better understanding of human infertility (Eberhart et al., 1996).
Boule gene was shown to be a key regulator of meiosis in Drosophila. Boule-deficient fruit flies are infertile and display meiotic arrest in their male germ cells (Eberhart et al., 1996 ). In fruit flies, boule is expressed in the prophase of the first meiotic division. It regulates the expression of Twine, Cdc25 phosphatase required to activate the maturation promoting factor, consisting of the Cdc2/cyclinB complex. It is crucial for entering the G2/M transition phase to complete the first meiotic division (Mains and Wasserman, 1999; Xu et al., 2003). Recently the human Boule gene might have the same or a very similar function as the Drosophila homolog as a regulator of meiosis because spermatogenesis was restored in boule mutant flies made transgenic for the human Boule gene (Xu et al., 2003). This suggests that the expression of Boule might be important for meiotic transition in the human as well.
The objectives of this study is to investigate the impact of some nitrogenous fertilizers on boule gene, isolate and characterize the boule gene in Drosophila melanogaster.
MATERIALS AND METHODS
This research had been performed in the Molecular Biology and Immunology Lab. In the Department of Entomology, Faculty of Science, Cairo University.
Drosophila stock: In the present study Drosophila stock was originally supplied by the Environmental Mutagens Research Unit (EMRU), Faculty of Agriculture, Ain Shams University. Drosophila was maintained on standard corn meal/agar/sucrose medium at 25°C. Wild-type flies were Oregon-R. (OR).
Tested chemicals: Nitrogenous fertilizers used are: Urea, Ammonium nitrate and Calcium nitrate.
Urea: CO(NH2)2 {Mol. Wt. 60.06}: Urea provided as a water-soluble white crystalline substance containing 46% nitrogen per g mol. Urea is converted to ammonia which reacts with water to form ammonium. Some volatilization of ammonia can occur when urea is surface applied.
Calcium nitrate: Ca (NO3)2 {Mol. Wt. 164}: The salt existed as water-soluble, pale brown crystalline solid with 16% nitrogen per g mol. The salt contains nitrogen in nitrate form.
Ammonium nitrate: NH4NO3 {Mol. Wt. 80.05}: It is a water soluble white crystalline substance containing 33% nitrogen per g mol. One-half of the nitrogen is in nitrate form which makes it immediately susceptible to potential leaching.
Larval treatment: The three nitrogenous compounds were administrated through larval feeding. For this purpose the tested substance was mixed throughout the regular medium just before it starts to solidify. Newly hatched larvae were put on the tested chemicals (LC25, LC50 determined before) and incubated at 25°C. The newly emerged adult males were used for molecular assays. This study had been performed in Cell Biology Department National Research Center, Egypt.
RNA isolation and RT. PCR analysis: was extracted from male adults using Gentra Purescript for RNA Kit (Life Trade company). Three micrograms of total RNA was used for RT-PCR reactions in 25 μL total volume using RobustTM II RT-PCR Kit (Robust and DyNAzyme are trademarks of Finnzymes Oy) with the following forward (bol. 1) and reverse (bol. 2) primers (designed according to Ayyar et al., 2003).
| bol. 1: AAACGCATCGTATCTGGG. bol. 2: TGAAGGTGGGTAGATGGC. |
The thermal cycle program was set to heat at 42°C for 60 min, at 94°C for 4 min, followed by 36 cycles of denaturation at 94°C for 1 min, 52°C annealing for 1 min and 72°C elongation for 2 min each step. Followed by 10 min final extension at 72°C, using Biometra Thermal cycler.
PCR products were recovered and sequenced (using bol. 1 and bol. 2 primers) by automated DNA sequencing reactions, which were performed using sequencing ready reaction big dye terminator kit (Applied Biosystems, USA) in conjunction with ABI-PRISM 310 genetic analyser. The sequences were provided by MWG Biotech (Ebersberg, Germany). Primers used for the sequencing of the PCR products were the same as for the amplification.
RESULTS AND DISCUSSION
The mutagenic effects of some nitrogenous fertilizers indicated a noticeable incidence of sterility in males as well as in females Drosophila melanogaster in postmeiotic and meiotic stages (Adel et al., 2000). Spermatogenesis is strikingly similar between Drosophila and mammals (Fuller, 1998). Transcriptional activation in spermatocytes furnishes material that supports spermatocyte maturation and meiosis as well as further spermatid differentiation. Execution of the meiosis differentiation program requires the meiotic arrest genes. The aly, can, mia and sa genes of Drosophila are essential in males both for the G2/meiosis I transition and for onset of spermatid differentiation. Functions of all four genes is required for transcription in primary spermatocytes of a suit of spermatide differentiation genes. Aly gene is also required for transcription of the cell cycle control genes cyclinB, boule and twine in primary spermatocytes. In contrast, can, mia and sa genes are required for accumulation of twine protein at the translation level. It is proposed that the can, mia and sa gene products act together or in a pathway to turn on transcription of spermatide differentiation genes and that aly gene acts upstream of can, mia and sa genes to regulate spermatid differentiation. The present study is focused upon the effect of nitrogenous fertilizers on the variation in genetic structure of the Drosophila boule gene. Spermatocytes are formed in boule mutants, but fail to undergo meiotic divisions. Comparison of the localization of Cyclin A protein in boule mutant and in wild type supports the conclusion that the meiotic prophase is normal in boule mutants. Although the meiotic prophase appears wild type in boule mutant germ cell, subsequent stages are aberrant. Cyclin A is exclusively cytoplasmic in the extended premeiotic G2 and M phases only to be degraded rapidly after nuclear translocation, yet Cyclin A persists in boule mutants (Eberhart, 1996). Mikhaylova et al. (2006) purified the protein that specifically binds to the promoter of spermatid-differentiation gene Sdic and identified it as Modulo, the Drosophila homologue of nucleolin. Analysis of the gene-expression patterns in the male sterile Modulo mutant indicates that Modulo supports high expression of the meiotic-arrest genes and is essential for transcription of spermatid-differentiation genes. Expression of Modulo itself is under the control of meiotic arrest genes and requires the DAZ/DAZL homologue boule that is involved in the control of G2/M transition. Boule regulates the G2/M transition in meiosis by positive translational regulation of Cdc25/Twine (Perezgasga et al., 2004), Modulo and the products of the meiotic-arrest genes are required for expression of a number of spermatid-differentiation genes. They also concluded that regulatory interactions among Modulo, boule and the meiotic arrest genes integrate meiosis and spermatid differentiation in the male germ line. Courtot et al. (1992) illustrated that Twine is expressed in the growing stage of primary spermatocytes in a manner that suggests a role in regulating the entry into meiosis and analysis of a Twine mutation has demonstrated a requirement for the gene not only in male, but also in female meiosis (Fig. 1a).
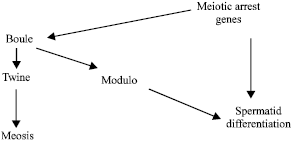 | |
| Fig. 1a: | Regulation of Modulo expression in testes by the meiotic-arrest genes and Boule links the pathways leading to meiosis spematid differentiation |
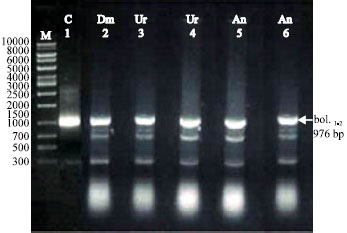 | |
| Fig. 1b: | RT-PCR analysis of total RNA prepared from adult male D. melanogaster (Dm). Lane M (DNA marker), Lane 1C (positive control), Lane 2 Dm (untreated), Lanes 3,4 (treated Dm with Urea (Ur) LC25 and LC50, respectively). Lanes 5,6 (treated Dm with Ammonium nitrate (An) LC25 and LC50, respectively). For amplification in lanes 2-6, as arrow indicates primers bol. 1 and bol. 2 were used and produce fragment of 976 bp |
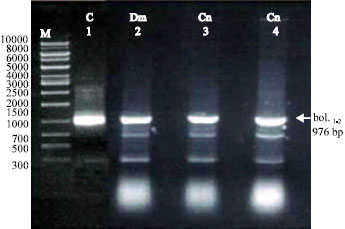 | |
| Fig. 2: | RT-PCR analysis of total RNA prepared from adult male D. melanogaster (Dm). Lane M (DNA marker), Lane 1 C (positive control), Lane 2 Dm (untreated), Lanes 3,4 (treated Dm with calcium nitrate (Cn) LC25 and LC50, respectively). For amplification in lanes 2-4, as arrow indicates primers bol. 1 and bol. 2 were used and produce fragment of 976 bp |
In present study, we performed RT-PCR analysis on total RNA extracted from untreated (control) and treated D. melanogaster adult male flies with LC25 and LC50 of the tested nitrogenous fertilizers Urea (Ur) and Ammonium Nitrate (An) (Fig. 1b) and with LC25 and LC50 of Calcium Nitrate (Cn) (Fig. 2). In both cases one pair of specific primer was used for amplification. The amplified products were found to be the same length of 976 bp in the untreated and treated adult male flies.
| Table 1: | The effect of LC25 and LC50 of the three tested nitrogenous fertilizers urea (Ur), Ammonium nitrate (An) and Calcium nitrate (Cn) on Drosophila melanogaster boule gene |
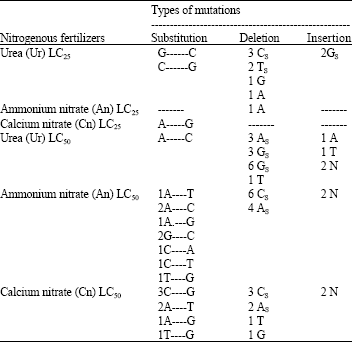 | |
| A means adenine, G means guanine, C means cytosine, T means thymine N undetectable nucleotide by the sequencer | |
The sequence of the boule gene fragment of the untreated adult males of the tested D. melanogaster Oregon-R showed 99% similarity at the nucleotide level to the corresponding boule gene fragment of D. melanogaster which has been submitted to the GenBank data library under accession no.U51858 (Fig. 3a and b). The sequence of the boule fragments of the treated adult male with LC25 of Ur, An and Cn, respectively showed different types of mutations include, substitution, deletion and insertion mutations. Ur induced substitution mutations G-C and C-G, deletion mutations and Gs insertion mutation. An induced deletion mutation. Cn induced substitution mutation A-G (Fig. 4 and Table 1).
The sequence of the boule fragments of the treated adult male with LC50 of Ur, An and Cn, respectively showed many mutations. Ur induced substitution mutations A-C, deletion and insertion mutations. Substitution mutations A-C, A-T, A-G, C-A, C-T, G-C,T-G and G-T, deletion and insertion mutations were recorded as an effect of An. Also, Cn induced substitution mutations T-A, T-C and G-C as well as deletion and insertion mutations (Fig. 5 and Table 1). We noticed that the diferrent types of mutations with LC50 of the tested nitrogenous fertilizers occur at the same region of the sequenced fragments. Tosal et al. (1998) showed that the most mutagenic sites in spermatogonial stem cells of Drosophila are A:T pairs (85%), with AT-->TA transversions (50%) and AT-->GC transitions (35%) as the most frequent mutations.
 | |
| Fig. 3a: | The sequence of boule gene of Drosophila melanogaster Oregon R. (Dm OR ) |
 | |
| Fig. 3b: | Alignment of the sequence of boule gene of the tested Drosophila melanogaster Oregon R and the sequence of Drosophila melanogaster boule gene which has been submitted to the GenBank data library under accession No. U51858.The non- bold letters indicate the differences between them |
Also, Petrov and Hartl (1999) concluded that the frequency of different nucleotide substitutions are far from equal. The greatest bias is in frequency of transition from G.C---A.T, which is on average 2.2 times more frequent than any other substitution. The other possible transition, A.T-G.C, is much less frequent and is, in fact, less frequent than some of the transversions. The transition bias in Drosophila therefore is a bias only toward the G.C-A.T transition.
Bentley et al. (2000) observed that the Ethyl-Methanesulfonate (EMS) mutational spectrum in Drosophila germ cells shows a strong preference for 5-UG-3 sites and for G/C within a stretch of three or more G/C base pairs. Also, Rincon et al. (1998) concluded that treatment of Drosophila larvae with the mixture resulting from the in vitro reaction of nitrosation precursors (methyl urea, sodium nitrite or its combination) resulted in high frequencies of induced wing spots comparable to those recorded with the potent genotoxin N-nitrosomethylurea. Also, Ryskova et al. (1997) demonstrated the genotoxicity of N-nitroso-N-methylurea (MNU) and Acetone Oxime (ACOX) in Drosophila melanogaster and determined the effect of human glutathione S-transferase on the genotoxic response. MNU was highly genotoxic in both transgenic flies {expressing the human gene encoding a glutathione S-transferase alpha subunit (HGST)} and non-transgenic flies. The non-transgenic flies were significantly more sensitive to the genotoxic effects of MNU compared to transgenic flies. ACOX also proved to be genotoxic to both non-transgenic and transgenic flies. While, Nevo and Coll (2001) reported that Aphid on nitrogen-fertilized plants were significant bigger and darker. All body size and darkness of color measurements were positively correlated with Aphid fecundity. Urea derived compounds such as: 1,4 Dioxane (DX) and Thiourea (TU) have the functional ability to work as rodent carcinogenesis on Drosophila. TU causes increase of meiotic Non-Disjunction (ND) in the earliest cells tested in a more consistent fashion than DX.
 | |
| Fig. 4: | Alignment of the sequence of untreated Drosophila melanogaster male boule gene (Dm) and treated with LC25 of the three tested nitrogenous fertilizers Urea (Ur), Ammonium Nitrate (An) and Calcium Nitrate (Cn), respectively dashes mean deletion |
 | |
| Fig. 5: | Alignment of the sequence of untreated Drosophila melanogaster male boule gene (Dm) and treated with LC50 of the three tested nitrogenous fertilizers Urea (Ur), Ammonium Nitrate (An) and Calcium Nitrate (Cn), respectively. Non-bold letters indicate the different types of mutation. N (undetectable nucleotide by the sequencer) and dashes mean deletion |
Toxicity of DX and TU is also known to play a significant role in the induction of chromosome malsegregation (Munoz et al., 2002). Baldrich et al. (2003) illustrated that the White-apricot (Wa) mutant of Drosophila melanogaster is characterized by a copia retrotransposon inserted in the second intron of the white locus. After germinal exposure to alkylating agent N-ethyl-N-nitrosourea, they obtained new phenotypes seem to be caused by mutations being induced in other loci acting as modifiers, most of them located on the X chromosome. Moreover, mutation frequencies induced by urea may be attributed to gene mutations rather than chromosomal aberrations since urea significant alternations were higher in the earlier stages of spermatogensis where DNA replication takes place. On the contrary the lethal mutation induced by ammonium nitrate and calcium nitrate in late spermatogenesis may be a consequence of both gene mutations as well as chromosomal aberration. Similar results were recorded by Lee et al. (1983).
The results obtained in this study indicate that the three nitrogenous fertilizers used possess mutagenic potentialities. The mutagenic activity is due to their Nitrogen (N) content. Another possible interpretation for the mutagenicity of the tested compounds is the ability of nitrogenous fertilizers to methylate DNA resulting in different types of genetic alterations (Colutier et al., 1999). Regard to the effect of nitrogenous fertilizers on the variation in genetic structure of the Drosophila boule gene, they induced different types of mutations in the boule gene such as substitution, deletion and insertion mutations. Mutation of boule gene blocks both meiotic divisions leading to tetraploid spermatids that fail to mature into spermatozoa. Boule mutants carry out chromosome condensation and centrosome duplication but are incapable of spindle formation, nuclear lamina breakdown, or chromosome disposition at the metaphase plate. Boule, is essential for the meiotic progression in spermatogenesis (Eberhart et al., 1996).
Wakimoto et al. (2004) performed a large-scale screen for male-sterile mutations. From a collection of 12, 326 strains carrying ethyl-methanesulfonate-treated, homozygous viable second or third chromosomes, 2216 male sterile lines were identified, constituting the largest collection of male sterile mutations described up to date for any organism. Over 2000 lines were cytologically characterized and, of these, 81% failed during spermatogenesis while, 19% manifested postspermatogenic processes. They also identified 62 fertile or subfertile lines that showed high levels of chromosome loss due to abnormal mitotic or meiotic chromosome transmission in the male germ line or due to paternal chromosome loss in the early embryo. They argued that the majority of autosomal genes that function in male fertility in Drosophila are represented by one or more alleles in the male sterile collection. Analysis of these mutations should provide insight into the genetic networks that control male fertility in Drosophila and other organisms, including humans.
Further studies are required to determine comparative structure and sequence, domain organization and valuable recognition motif in the coding region of boule, DAZ (human) and Dazla (mouse) and similar functional protein in the process of spermatogenesis. Sequencing and comparative studies of the impact of different mutation on the coding region as a termination signals for these proteins are important for investigating the mode of action of fertility and sterility in respond to mutation for boule gene and other similar fertility genes.
REFERENCES
- Alvarez, L., M.A. Comendador and L.M. Sierra, 2002. O-ethylthymidine adducts are the most relevant damages for mutation induced by N-ethyl-N-nitrosourea in female germ cells of Drosophila melanogaster. Environ. Mol. Mutagen, 40: 143-152.
Direct Link - Ayyar, S., J. Jiang, A. Collu, H. White-Cooper and A.H. White Robert, 2003. Drosophila TGIF is essential for developmentally regulated transcription in spermatogenesis. Development, 130: 2841-2852.
CrossRefDirect Link - Baldrich, E., A. Velazquez, N. Xamena and O. Carbre, 2003. Germline mutations induced by N-nitroso-N-ethylurea do not affect the inserted copia retrotransposon in a Drosophila melanogaster wa mutant. Mutagenesis, 18: 527-531.
Direct Link - Bentley, A., B. McLennan, J. Calvo and C.R. Dearolf, 2000. Targeted recovery of mutations in Drosophila. Genetics, 156: 1169-1173.
PubMedDirect Link - Colutier, J.F., R. Drouin and A. Castonguay, 1999. Treatment of human cells with N-nitroso (acetoxymethyl) methylamine: Distribution patterns of piperidine-sensitive DNA damage at the nucleotide level of resolution are related to sequence context. Chem. Res. Toxicol., 12: 840-849.
CrossRefDirect Link - David, C.L., V.A. Pierce, D.W. Aswad and A.G. Gibbs, 1999. The effect of urea exposure on isoaspartyl content and protein L-isoaspartate methyltransferase activity in Drosophila melanogaster. Comp. Biochem. Physiol. Biochem. Mol. Biol., Dec., 124: 423-427.
CrossRefDirect Link - Eberhart, C.G., J.Z. Maines and S.A. Wasserman, 1996. Meiotic cell cycle requirement for a fly homologue of human deleted in Azoospermia. Nature, 381: 783-785.
CrossRefDirect Link - Fuller, M.T., 1998. Genetic control of cell proliferation and differentiation in Drosophila spermatogenesis. Semin. Cell. Dev. Biol., 9: 433-444.
Direct Link - Mains, J.Z. and S.A. Wasserman, 1999. Post-transcriptional regulation of the meiotic Cdc25 protein Twine by the Dazl orthologue Boule. Nat. Cell Biol., 1: 171-174.
CrossRefDirect Link - Mensinga, T.T., G.J. Spijers and J. Meulenbelt, 2003. Health implications of exposure to enviromental nitrogenous compounds. Toxicol. Rev., 22: 41-51.
PubMedDirect Link - Miasoedova, E.E. and S.B. Nazarov, 2004. The response of erythrocytic system of mature rats to acute nitrite intoxication. Patol. Fiziol. Eksp. Ter., 2: 16-18.
PubMedDirect Link - Mikhaylova, L.M., A.M. Boutanaev and D.I. Nurminsky, 2006. Transcriptional regulation by Modulo integrates meiosis and spermatid differentiation in male germ line. PNAS, 103: 11975-11980.
Direct Link - National Toxicology Program, 2001. Toxicology and carcinogenesis studies of sodium nitrite CAS No. 7632-00-0 in F344/N rats and B6C3F1 mice drinking water studies. Nat. Toxicol. Program Tech. Rep. Ser., 495: 7-273.
PubMedDirect Link - Ohsawa, K., S.Y. Nakagawa, M. Kimura, C. Shimada and S. Tsuda et al., 2003. Detection of in vivo genotoxicity of endogenously formed N-nitroso compounds and suppression by ascorbic acid, teas and fruit juices. Mut. Res. Aug., 539: 65-76.
Direct Link - Perezgasga, L., J. Jiang, B. Jr. Bolival, M. Hiller, E. Benson, M.T. Fuller and H. White-Cooper, 2004. Regulation of transcription of meiotic cell cycle and terminal differentiation genes by the testis-specific Zn-finger protein matotopetli. Development, 131: 1691-1702.
PubMedDirect Link - Petrov, D.A. and D.L. Hartl, 1999. Patterns of nucleotide substitution in Drosophila and man. Proc. Nat. Acad Sci. USA., 96: 1475-1479.
Direct Link - Rincon, G.J., J. Espinosa and U. Graf, 1998. Analysis of the in vivo nitrosation capacity of the larvae used in the wing somatic mutation and recombination test of Drosophila melanogaster. Mut. Res., 412: 69-81.
Direct Link - Ryskova, M., K. Chroust, M. Trbusek, J. Benedik and T. Jowett, 1997. Genotoxicity of N-nitroso-N-methylurea and acetone oxime in the transgenic Drosophila carrying the human gene encoding a subunit of glutathione S-transferase. Folia Biol. Praha, 43: 19-24.
PubMedDirect Link - Tosal, L., M.A. Comendador and L.M. Sierra, 1998. N-ethyl-N-nitrosourea predominantly induces mutations at A T base pairs in pre-meiotic germ cells of Drosophila males. Mutagenesis, 13: 375-380.
Direct Link - Wakimoto, B.T., D.L. Lindsley and C. Herrera, 2004. Toward a comprehensive genetic analysis of male fertility in Drosophila melanogaster. Genetics, 167: 207-216.
Direct Link - White-Cooper, H., D. Leroy, A. MacQueen and M.T. Fuller, 2000. Development transcription of meiotic cell cycle and terminal differentiation genes depends on a conserved chromatin associated protein, whose nuclear localization is regulated. Development, 127: 5463-5473.
Direct Link