
Abdulkhaliq A. AL-Shoaibi
Department of Biology, Faculty of Science, Taibah University, P.O. Box 30002, Almadinah Almunawwarah, KSA
Journal of Biological Sciences
Year: 2008 | Volume: 8 | Issue: 3 | Page No.: 610-615







ABSTRACT
Photosynthetic responses of the C4 grass (Pennisetum purpureum) to three NaCl salinity concentrations (100, 200 and 400 mM) along with control (0 mM NaCl) were studied. The results showed that all the three salinity treatments did not affect the quantum efficiency of PSII photochemistry (Fv/Fm) compared to control. However, only the highest salinity concentration of 400 mM, significantly reduced (p<0.001) the light saturated assimilation rate (Asat), the quantum yield (φ), the CO2 saturated rate of CO2 uptake (Amax) and the carboxylation efficiency of P. purpureum, compared to control. Therefore, it is possible for P. purpureum to attain high rates of photosynthesis under low and medium salinity levels.
PDF Abstract XML References Citation
How to cite this article
Abdulkhaliq A. AL-Shoaibi, 2008. Photosynthetic Response of Elephant Grass (Pennisetum purpureum) to NaCl Salinity. Journal of Biological Sciences, 8: 610-615.
DOI: 10.3923/jbs.2008.610.615
URL: https://scialert.net/abstract/?doi=jbs.2008.610.615
DOI: 10.3923/jbs.2008.610.615
URL: https://scialert.net/abstract/?doi=jbs.2008.610.615
INTRODUCTION
The salinity is considered as one of the major causes that influence the photosynthetic efficiency of plants either in the controlled environments or in the field (Munns et al., 2000; Gibberd et al., 2002; Blindow et al., 2003; Lycoskoufis et al., 2005; Youssef, 2007). Most of the commercially important crops are salt sensitive and as a consequence of the salinity may limit their growth and production (Flowers, 2004). A previous study of photosynthetic gas exchange in the C4 plant Atriplex lentiformis (Torr.) showed high rates of photosynthesis could be attained at low levels of NaCl salinity (Zhu and Meinzer, 1999). In addition, the high levels of NaCl salinity impaired the CO2 assimilation in this C4 plant. The inhibition of photosynthesis by high NaCl concentration is usually associated with the reduction of both the light-saturated rate of carbon dioxide assimilation (Asat) and the maximum quantum (φ) yield (Lycoskoufis et al., 2005; Lopez-Climent et al., 2008; Youssef, 2007).
The phenomenon of photoinhibition is typically manifested as a light-dependent decrease in the quantum yield of photosystem II (PSII) photochemistry (Hu et al., 2006). The linear relationship between quantum yield and the ratio of variable fluorescence to maximum fluorescence (Fv/Fm) (Adams et al., 1990) indicate that Fv/Fm can monitor the photosynthetic carbon assimilation (Björkman, 1987). The combination of chlorophyll fluorescence measurement together with the net gas exchange parameters provides a good way of the evaluation of the photosynthetic performance in the stressed plants (Jimenez et al., 1997) and to gain an insight into the behavior of the photosynthetic machinery under such stress (Maxwell and Johnson, 2000). Several plants have been subjected to photoinhibition at salinity treatment, but others showed no change in the Fv/Fm values among salinity treatments (Arfan et al., 2007; Broetto et al., 2007; Lopez-Climent et al., 2008; Neocleous and Vasilakakis, 2007).
Elephant grass (P. purpureum) is a fast-growing C4 grass (Wang et al., 2002) cultivated for cattle feeding in the Kingdom of Saudi Arabia. Salinity is the maim concern for the potential commercial production of elephant grass in the arid regions of the world (Wang et al., 2002). The previous results of Al-Shoaibi and Al-Sobhi, (2004) confirmed that the salinity arising from irrigation caused a major problem for P. purpureum growth in Al-Madinah Al-Munawwarah in the western part of the Kingdom of Saudi Arabia. This lower growth rate at high salinity levels could be as a result of the reduction in the photosynthetic capacity. Since there is a lack of the knowledge via published reports about the influence of salinity on gas exchange of P. purpureum, this study was carried out to demonstrate the response of the photosynthetic CO2 uptake of P. purpureum to NaCl salinity.
MATERIALS AND METHODS
Plant and growth conditions: This research was conducted in Biology Department, Faculty of Science, Taibah University during 2006. Rhizomes of Pennisetum purpureum originally derived from Africa were planted in 12 L pots filled with coarse sand and grown in high-light, in a controlled environment chamber (Fitotron SGC066. CHX, Sanyo Gallenkamp PLC, Leicester, UK.), at day/night temperatures of about 25/20 °C. The VPD was kept below 1 kPa. Photon flux density at leaf height was 600 μmol m-2sec-1 and the photo period was 14 h.
Three NaCl concentrations, 100, 200 and 400 mM, in full strength Hoagland nutrient solution were used (Arnon and Hoagland, 1940). The control was Hoagland nutrient solution without addition of NaCl. All pots were irrigated every other day for 7 days with full strength Hoagland nutrient solution. NaCl treatments begun 9 days after the start of the experiment. The NaCl concentration was increased in steps of 100 mM on alternate days until the appropriate salt treatments were reached.
Gas exchange and chlorophyll fluorescence measurements: The CO2 uptake was measured using an open gas-exchange system incorporating open path infrared, CO2 and water vapour analysers (LI-6400, LI-COR Inc., Lincoln, USA). A Peltier cooling system maintained the leaf temperature at 20 °C as described by Long et al. (1996) and VPD was controlled between 1 and 1.5 kPa. The most recent fully expanded leaf was used to measure photosynthetic CO2 uptake at PPFD of 0-2000 μmol m-2 sec-1. Net photosynthesis per unit leaf area and intercellular CO2 concentration (ci) were determined using the equations of Von Caemmerer and Farquhar (1981). The light saturated photosynthesis (Asat) was determined at saturated PPFD (1500 μmol m-2 sec-1) and at the ambient CO2 concentration of 360 μmol mol-1. As well, The light-limited photosynthetic capacity was measured at the ambient CO2 concentration by increasing PPFD in three steps to 150 μmol m-2 sec-1 (0-50, 50-100 and 100-150 μmol m-2 sec-1). Carbon dioxide response curves were made over the range of 50-550 μmol mol-1 using PPFD of 1500 μmol m-2 sec-1, at a leaf temperature of 20 °C. These curves were analysed according to the model of Collatz et al. (1992).
Chlorophyll fluorescence was measured using a portable fluorimeter (PEA, Hansatech, Kings Lynn, Norfolk). The initial (F0) and maximum (Fm) fluorescence emissions were measured after 20 min of dark adaptation and the ratio of variable to maximum fluorescence (Fv/Fm) was calculated as described by Öquist and Wass (1988) from the following equation: Fm-F0/Fm. The Fv/Fm ratio was measured weekly for 4 replicates of each treatment, using the youngest fully expanded leaves.
Statistical analyses: One-way analysis of variance was used to study the effect of salinity on Fv/Fm, Asat, φ, Amax and the carboxylation efficiency of P. purpureum. Post hoc pairwise comparisons by Scheffe`s test were performed to detect differences between individual means (Systat, Inc., Evanston, Illinois. USA). The effect was described as significant where p<0.05.
RESULTS
The values of Fv/Fm were not significantly affected by the three salinity treatments compared to control. They always remained above 0.8 (Fig. 1).
The results shown in Fig. 2 revealed the response of photosynthetic CO2 uptake (A) to photon flux (Q) for P. purpureum at different salinity levels. The light-saturated (Asat) of P. purpureum leaves grown at 400 mM NaCl concentration was significantly decreased (p<0.001), compared to leaves grown control (Fig. 2). The reduction percentage was 36%, compared to the control reading. Also, the highest salinity concentration significantly reduced (p<0.001) the light-limited photosynthetic capacity (φ) of P. purpureum leaves, compared to leaves grown at control (Fig. 3). The reduction percentage was 21%, compared to control record. On the other hand, the Asat and φ of P. purpureum leaves grown at 100 and 200 mM NaCl were similar to those grown at control (Fig. 2-3).
The saturated rate of CO2 uptake (Amax) is co-limited by the amount of in vivo Rubisco or/and PPDK activities (Collatz et al., 1992). The results (Fig. 5) show that Amax was significantly reduced (p<0.001) at the highest level of
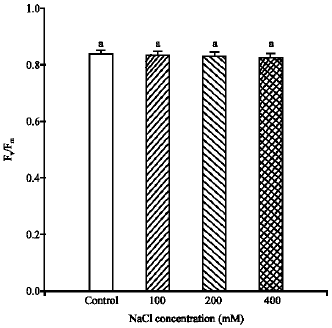 | |
| Fig. 1: | The dark adapted Fv/Fm of Pennisetum purpureum. The data represent the mean of n = 3-6 replicate shoots ( ± SE). Different letters show the significant differences between NaCl treatments at p<0.05 |
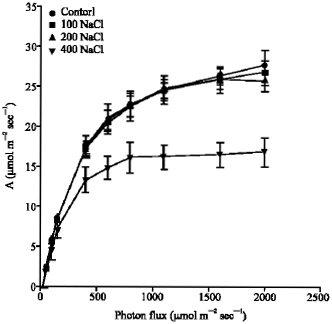 | |
| Fig. 2: | The response of photosynthetic CO2 uptake (A), per unit leaf area, to photon flux (Q) for Pennisetum purpureum. Measurements of CO2 uptake were all made at 20 °C and Ca of 360 μmol mol-1. The data represent the mean of n = 3-6 leaves ( ± SE) |
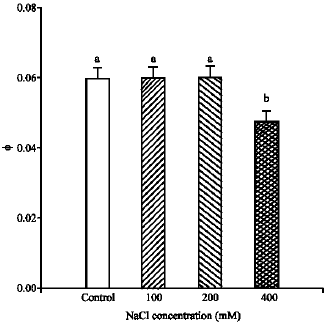 | |
| Fig. 3: | The mean quantum yield (φ), measured at 20 °C, for Pennisetum purpureum. The data represent the mean of n = 3-6 leaves ( ± SE). Different letters show the significant differences between NaCl treatment at p<0.05 |
salinity compared to control. The percent of reduction was 40% compared to control (Fig. 4-5). The Amax was not significantly decreased by 100 and 200 mM NaCl salinity
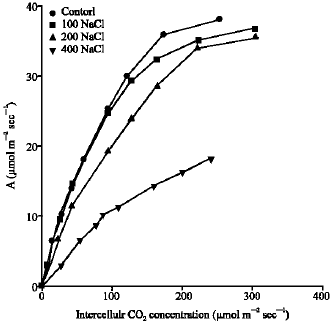 | |
| Fig. 4: | The response of photosynthetic CO2 uptake (A), per unit leaf area, to the changes in intercellular CO2 concentration (ci) for Pennisetum purpureum leaves. Measurements of CO2 uptake were all made at 20 °C and photon flux of 1500 μmol mol-1. Data illustrated are for one selected leaf of each plant |
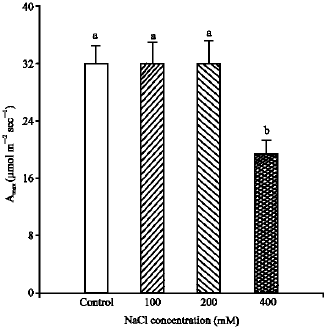 | |
| Fig. 5: | The mean plateau (Amax) per unit leaf area, measured at 20 °C and photon flux of 1500 μmol m-2 sec-1 for Pennisetum purpureum. The data represent the mean of n = 3-6 leaves ( ± SE). Different letter(s) show the significant differences between NaCl treatments at p<0.05 |
compared to control (Fig. 4-5). In addition, growth at the highest salinity treatment, in comparison to control, had
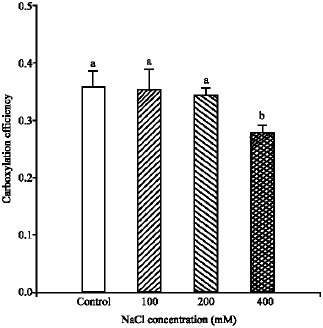 | |
| Fig. 6: | The mean carboxylation efficiency, measured at 20 °C and photon flux of 1500 μmol m-2 sec-1 for Pennisetum purpureum. The data represent the mean of n = 3-6 leaves ( ± SE). Different letter(s) show the significant differences between NaCl treatments at p<0.05 |
a significant affect (p<0.001) on the carboxylation efficiency of P. purpureum leaves (Fig. 6). The percentage of reduction was about 22%, compared to control. On the contrary, the carboxylation efficiency of P. purpureum leaves grown at 100 and 200 mM NaCl salinity were quite similar to those grown at the control (Fig. 6).
DISCUSSION
The photosynthetic response of P. purpureum to the salinity was investigated under salinity stress conditions. The results obtained in the present study showed that the quantum yield of PSII (Fv/Fm) was not significantly changed due to salt stress applications. The values of Fv/Fm always remained above 0.8 and there was no acute photoinhibition (Broetto et al., 2007). These high values of Fv/Fm provided a clear evidence of P. purpureum resistance to the photinhibition under different salinity treatments (Fig. 1). Photoinhibition occurs when the leaves are exposed to irradiances in excess of what can be utilized in photosynthesis (Powles, 1984). Similar results were previously reported for two wheat cultivars differing in their salinity tolerance (Arfan et al., 2007). Thus, this result suggests that the quantum yield of PSII (Fv/Fm) cannot be considered as one of the factors to regulate photosynthesis in P. purpureum under salinity treatments.
When P. purpureum grown under 100, 200 mM NaCl, all leaves showed similar Asat and φ to control (Fig. 2-3). The rates of Asat and φ were close to that recorded previously for healthy leaves of a range of NADP-malic enzymes type C4 grasses (Ehleringer and Pearcy, 1983). On the other hand, the 400 mM NaCl significantly reduce the Asat and φ of P. purpureum. However, rates of Asat and φ of P. purpureum grown at 400 mM NaCl were in excess of Zea mays grown at 200 mM of NaCl (Data not shown). This depression in the photosynthetic performance of P. purpureum could be due to the stomatal conductance or ion toxicity or both (Ashraf, 2004; Sudhir and Murthy, 2004). Another possibility is that the significant reduction in the photosynthetic capacity of P. purpureum at the highest concentration of salinity could be a result of reduction in the leaf area (Munns et al., 2000).
Analysis of A/ci curves indicated that the 400 mM NaCl treatment caused a significant reduction in Amax and carboxylation efficiency of P. purpureum (Fig. 4-6). Similar results were recorded for three wild soybean species regarding the difference in their response to NaCl treatments (Crosbie and Pearce, 1982; Kao et al., 2006) in response to salinity. The Amax and carboxylation efficiency are controlled by activity of Ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) and activity of Phosphoenol pyruvate carboxylase enzymes (PEPC), respectively (Collatz et al., 1992). Therefore, the inhibition of Amax by high NaCl salinity may cause a reduced efficiency of ribulose-1,5-bisphosphate (RuBP) carboxylase, or a reduction of RuBP regeneration capacity, or the sensitivity of PSII to NaCl concentration (Ball and Anderson, 1986). On the other hand, the reduced efficiency of PEPC enzyme may cause the inhibition of carboxylation efficiency by high NaCl salinity. In contrast, the rates of Amax and carboxylation efficiency of P. purpureum grown at 100 and 200 mM NaCl were similar to control. This may suggest that P. purpureum contains similar levels of Rubisco and PEPC regardless whether the growth at low or medium salinity.
In conclusion, the photosynthetic response of P. purpureum to the salinity was investigated under salinity stress conditions. The values of Fv/Fm for P. purpureum leaves grown at 100, 200 and 400 mM NaCl were similar to control and always remained above 0.8. Only the highest salinity concentration of 400 mM, significantly reduced the light saturated assimilation rate (Asat), the quantum yield (φ), the CO2 saturated rate of CO2 uptake (Amax) and the carboxylation efficiency of P. purpureum, compared to control. Therefore, P. purpureum can attain high photosynthetic rates under low and medium salinity levels.
ACKNOWLEDGMENT
The author is indebted to Professor Dr. Moustafa EL-Naggar for his critical reading and revision of the manuscript.
REFERENCES
- Adams, W.W., B. Demmig-Adams, K. Winter and U. Schreiber, 1990. The ratio of variable to maximum chlorophyll fluorescence from photosystem II. Measured in leaves at ambient temperature and at 77K, as an indicator of the photon yield of photosynthesis. Planta, 180: 166-174.
CrossRefDirect Link - Arfan, M., H.R. Athar and M. Ashraf, 2007. Does exogenous application of salicylic acid through the rooting medium modulate growth and photosynthetic capacity in two differently adapted spring wheat cultivars under salt stress?. J. Plant Physiol., 164: 685-694.
CrossRefPubMedDirect Link - Ashraf, M., 2004. Some important physiological selection criteria for salt tolerance in plants. Flora-Morphol. Distrib. Funct. Ecol. Plants, 199: 361-376.
CrossRefDirect Link - Ball, M.C. and J.M. Anderson, 1986. Sensitivity of photosystems II to NaCl in relation to salinity tolerance. Comparative studies with thylakoids of the salt tolerant mangrove, Avicennia marina and the salt-sensitive pea, Pisum sativum. Aust. J. Plant Physiol., 13: 689-698.
CrossRefDirect Link - Blindow, I., J. Dietrich, N. Möllmann and H. Schubert, 2003. Growth, photosynthesis and fertility of Chara aspera under different light and salinity conditions. Aquatic Bot., 76: 213-234.
Direct Link - Broetto, F., H.M. Duarteb and U. Luttgeb, 2007. Responses of chlorophyll fluorescence parameters of the facultative halophyte and C3-CAM intermediate species Mesembryanthemum crystallinum to salinity and high irradiance stress. J. Plant Physiol., 164: 904-912.
Direct Link - Crosbie, T.M. and R.B. Pearce, 1982. Effects of recurrent phenotypic selection for high and low photosynthesis on agronomic traits in two maize populations. Crop Sci., 22: 809-813.
CrossRefDirect Link - Ehleringer, J. and R.W. Pearcy, 1983. Variation in quantum yield for CO2 uptake among C3 and C4 plants. Plant Physiol., 73: 555-559.
Direct Link - Gibberd, M.R., N.C. Turner and R. Storey, 2002. Influence of saline irrigation on growth, ion accumulation and partitioning and leaf gas exchange of carrot (Daucus carota L.). Ann. Bot., 90: 715-724.
Direct Link - Hu, H.W., Y.H. Zhou, Y.S. Du, X.J. Xia and J.Q. Yu, 2006. Differential response of photosynthesis in greenhouse- and field-ecotypes of tomato to long-term chilling under low light. J. Plant Physiol., 163: 1238-1246.
CrossRefPubMedDirect Link - Jimenez, M.S., A.M. Gonzalez-Rodriguez, D. Morales, M.C. Cid, A.R. Socorro and M. Caballero, 1997. Evaluation of chlorophyll fluorescence as a tool for salt stress detection in roses. Photosynthetica, 33: 291-301.
CrossRef - Kao, W.Y., T.T. Tsai, H.C. Tsai and C.N. Shih, 2006. Response of three Glycine species to salt stress. Environ. Exp. Bot., 56: 120-125.
Direct Link - Lopez-Climent, M., V. Arbona, R.M. Perez-Clemente and A. Gomez-Cadenas, 2008. Relationship between salt tolerance and photosynthetic machinery performance in citrus. Environ. Exp. Bot., 62: 176-184.
Direct Link - Long, S.P., P.K. Farage and R.L. Garcia, 1996. Measurements of leaf and canopy photosynthesis CO2 exchange in the field. J. Exp. Bot., 47: 1629-1642.
Direct Link - Lycoskoufis, I.H., D. Savvas and G. Mavrogianopoulos, 2005. Growth, gas exchange and nutrient status in pepper (Capsicum annuum L.) grown in recirculating nutrient solution as affected by salinity imposed to half of the root system. Scientia Hortic., 106: 147-161.
Direct Link - Maxwell, K. and G.N. Johnson, 2000. Chlorophyll fluorescence-A practical guide. J. Exp. Bot., 51: 659-668.
CrossRefDirect Link - Munns, R., R.A. Hare, R.A. James and G.J. Rebetzke, 2000. Genetic variation for salt tolerance of durum wheat. Aust. J. Agric. Res., 51: 69-74.
Direct Link - Neocleous, D. and M. Vasilakakis, 2007. Effects of NaCl stress on red raspberry (Rubus idaeus L. Autumn Bliss). Scientia Hortic., 112: 282-289.
CrossRefDirect Link - Oquist, G. and R. Wass, 1988. A portable microprocessor operated instrument for measuring chlorophyll fluorescence kinetics in stress physiology. Physiol. Plant., 73: 211-217.
CrossRefDirect Link - Powles, S.B., 1984. Photoinhibition of photosynthesis induced by visible light. Annu. Rev. Plant Physiol., 35: 15-44.
CrossRefDirect Link - Sudhir, P. and S.D.S. Murthy, 2004. Effects of salt stress on basic processes of photosynthesis. Photosynthetica, 42: 481-486.
CrossRefDirect Link - Von Caemmerer, S. and G.D. Farquhar, 1981. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta, 153: 376-387.
CrossRefDirect Link - Wang, D., J.A. Poss, T.J. Donovan, M.C. Shannon and S.M. Lesch, 2002. Biophysical properties and biomass production of elephant grass under saline conditions. J. Arid Environ., 52: 447-456.
CrossRefDirect Link - Youssef, T., 2007. Stomatal, biochemical and morphological factors limiting photosynthetic gas exchange in the mangrove associate Hibiscus tiliaceus under saline and arid environment. Aquat. Bot., 87: 292-298.
Direct Link - Zhu, J. and C.F. Meinzer, 1999. Efficiency of C4 photosynthesis in Atriplex lentiformis under salinity stress. Aust. J. Plant Physiol., 26: 79-86.
CrossRef