
A. G. Pillay
Department of Anatomy, Faculty of Medicine, University of Malaya 50603, Kuala Lumpur, Malaysia
Pakistan Journal of Biological Sciences
Year: 2000 | Volume: 3 | Issue: 1 | Page No.: 52-56







ABSTRACT
Chick embryos at different stages of development till hatching as well as vestibular ganglia of the adult were fixed in formol-saline. Serial sections were stained by Cresyl Fast Violet to analyse the results in the vestibular ganglion. Neurones were categorised and counted. The appearance of a few light cells on Embryonic day 6 (E6) might indirectly indicate the beginning of an early establishment of functional connections in the vestibular ganglion in contrast to most other ganglia studied. The fluctuation in the number of cells during embryonic development may be considered as a normal process for the purpose of re-arrangement for better organisation in order to perform an efficient function. Volume of the vestibular ganglion is the greatest on Embryonic-day 18 (E18) during the whole ontogeny even though its rostro-caudal length is greatest in the adult. This is in contrast to that observed in several other ganglia where the ganglionic volume is the greatest in the adult situation. The reduction or loss of ganglionic neurones in the adult in comparison to that observed on the day of hatching might indicate a functional reduction as a result of ageing process. The appearance of an increased number of tiny cells in the adult in comparison to that observed on the day of hatching is different and peculiar from that observed in other ganglia studied. These new cells are probably derived from active mitoses of reserved neuroblasts. This possibly provides an attempt to replace the inactive / or dead cells during ageing process.
PDF Abstract XML References Citation
How to cite this article
A. G. Pillay, 2000. Vestibular Ganglion as a Model System of Vital-neural Centre During Embryonic Development. Pakistan Journal of Biological Sciences, 3: 52-56.
DOI: 10.3923/pjbs.2000.52.56
URL: https://scialert.net/abstract/?doi=pjbs.2000.52.56
DOI: 10.3923/pjbs.2000.52.56
URL: https://scialert.net/abstract/?doi=pjbs.2000.52.56
INTRODUCTION
During development and growth, the ganglionic volume and its cellular packing density vary greatly and they do not correspond with each other. On the contrary, in some stages during development there is increased ganglionic volume while there is reduction in the number of cells. In other, periods while there is reduced size of the ganglion, the cellular density is increased. Still in some other stages, there is increase both in ganglionic volume and cellular packing density. The onset of first-appearance of the dark and light cells in the ganglion and the fluctuation in their number and distribution pose great challenge in understanding the behavior of these cells. Interpretations of the significance of these cells vary among different authors (Gaik and Farbman, 1973; Spassova, 1982). More-over, most studies found in the literature. (Gaik and Farbman, 1973) are performed only in certain stages of development or growth that do not give a clear picture of the significance of such variation. Therefore, this study is aimed at analysing the cause of such fluctuation in the whole life of a single animal species. The animal model selected is the chick, Gallus gallus domesticus.
MATERIALS AND METHODS
The chicks Gallus gallus domesticus, White Leghorn. breed were used in this study. Fertilised eggs were incubated. After every 24 hours, it was considered as Embryonic Day 1 (E1), Embryonic Day 2 (E2) etc till hatching (H). Embryos till hatching were removed carefully and fixed in 10 percent formaldehyde solution at least for two weeks. Larger (older) embryos were cut transversely into suitable smaller pieces and labelled serially for future orientation. The tissues of older embryos (i.e., E15 and onwards till adult) were usually decalcified after fixation. Serial sections of 8-10 microns were stained by Cresyl Fast Violet for Nissl granules. Only a few selected stages that showed some remarkable changes are described in this work. These include E6, E8, E10, E13, E15, E18, chicks on the day of hatching (H) and adult (A). In all, three animals in each group, having a total of twenty-four animals were used. Ganglia of both sides in each animal (i.e., 6 ganglia for each stage) and therefore 48 ganglia for the whole study were used for observation. Every section of the ganglion was observed and drawn. The cells are plotted in a diagram with the help of a light microscope having a camera lucida attachment. Different categories of neurones were classified into dark and light neurones based on the difference in the intensity of cytoplasmic stain. For a beginner, the difference in the intensity of cytoplasmic stain to distinguish the dark and light cells might be confusing. However, this difficulty is over-come by practice. Each of these types is again subdivided into various subclasses represented in the diagram by a symbol. Only those cells having a clear nucleus and a nucleolus were counted and measured with the help of an eye-piece graticule. The dimension of every cell was determined by calculating the average of the two measurements: one measurement taken on its long axis at the place where the nucleus and its nucleolus were very clearly observed and another taken at an angle perpendicular to this long axis. However, the possible error in calculating the size of the cells is considered to be very minimal or negligible. The following categories of cells were classified. Tiny (<5 microns), very small (6-10 microns), small (11-15 microns), medium sized (16-20 microns), big (21-25 microns), very big (26-30 microns) and large (31-35 microns). The categorisation of cells on the basis of size with a uniform difference of 5 microns was initially maintained just for the sake of convenience. However, this proved to be very useful in that, the behavior of cells, especially that of the very-small cells, is very interesting on the day of hatching (uniformly) in all the ganglia studied. This explains that this particular stage of cellular growth (very-small cell stage) is a critical period during development, indicating a stage of active cell-process-formation (axon-formation) beginning to establish functional connections with the target tissues. This information is also used to interpret the functional significance of the occurrence of light cells during early stages of development.
RESULTS
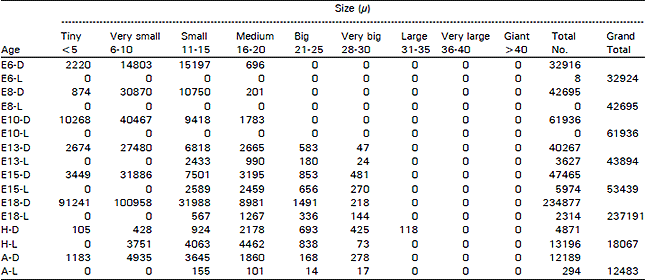
The ganglion showed great difference in different age groups of animals (E6 to E18, H, A) and in different areas in the same ganglion. The most striking changes are as follows. When the dark neurones alone are present, they are represented just by their numbers, when they are mixed with light cells, D= dark neurones and L= light neurones. Population density (P.D = in mm3) for certain types of cells is given in parenthesis (Fig. 1).
The vestibular ganglion could be recognised on E6 while it had a rostra-caudal length of 0.304 mm and a volume of 0.0094 mm3 and 32924 cells (P.D = 3502553). Among these cells, 32916 (99.98%) were dark type (P.D = 3501702) and 8 (0.02%) were light ones (P.D = 851). In all, there were 2220 (6.74%) tiny cells (P.D = 236170), 14803 (44.96%) very small ones (P.D = 1574787), 15197 (46.16%) small ones (greatest P.D = 1616702) and 704 (2.14%) medium sized ones (P.D = 74893). On ES, the ganglion had a length of 0.480 mm and a volume of 0.0240 mm3 and contained 42695 cells (P.D = 1778958) and all of these cells were dark type. In all, there were 874 (2.05%) tiny cells (P.D= 36417), 30870 (72.3%) very small ones (greatest P.D = 1286250), 10750 (25.18%) small ones (P.D = 447917) and 201 (0.47%) medium sized ones (P.D = 8375). On E10, the ganglion had a length of 0.963 mm, a volume of 0.0442 mm3 and 61936 cells (P.D = 1401267) and all of these cells were dark type. In all, there were 10268 (16.58%) tiny cells (P.D = 232308), 40467 (65.34%) very small ones (greatest P.D = 915543), 9418 (15.21%) small ones (P.D = 213077) and 1783 (2.88%) medium sized ones. On E13 3, the ganglion had a length of 1.020 mm, a volume of 0.1232 mm3 and 43894 cells (P.D = 356282). Among these cells, 40267 (91.74%) were dark type (P.D= 326842) and 3627 (8.26%) were light ones (P.D = 29440). In all, there were 2674 (6.09%) tiny cells (P.D = 21705), 27480 (62.61%) were very small ones (greatest P.D= 223052), 9251 (D= 6818 + L= 2433) (21.08%) were small ones (P.D = 75089), 3655 (D = 2665 + L = 990) (8.33%) were medium sized ones (P.D = 29667), 763 (0 = 583 + L = 180) (1.74%) w ere big ones (P.D=6193) and 71 (D = 47 + L = 24) (0.16%) were very big ones (P.D = 576). On E15, the ganglion had a length of 0.940 mm, a volume of 0.0770 mm3 and 53439 cells (P.D = 694013). Among these cells, 47465 (88.82%) were dark type (P.D = 616429) and 5974 (11.18%) were light ones (P.D= 77584). In all, there were 3449 (6.45%) tiny cells (P.D = 44792), 31886 (59.67%) were very small ones (greatest P.D = 414104), 10090 (D = 7501 + L= 2589) (18.88%) were small ones (P.D = 131039), 5654 (D = 3195 + L = 2459) (10.58%) were medium sized ones (P.D = 73429), 1609 (D = 953 + L= 656) (3.01% were big ones (P.D = 20896) and 751 (D = 481 + L = 270) (1.41%) were very big ones (P.D = 9753). On E18, the ganglion had a length of 1.230 mm, a volume of 0.3874 mm3 and 237191 cells (P.D = 612264). Among these cells, 234877 (99.02%) were dark type (P.D = 606291) and 2314 (0.98%) were light ones (P.D = 5973). In all, there were 91241 (38.47%) tiny cells (P.D = 235521), 100958 (42.56%) were very small ones (greatest P.D = 260604), 32555 (D = 31988 + L= 567) (13.73%) were small ones (P.D = 84035), 10248 (D = 8981 + L = 1267) (4.32%) were medium sized ones (P.D= 26453), 1827 (D= 1491 + L= 336) (0.77%) were big ones (P.D = 4716) and 362 (D = 218 + L= 144) (0.15%) were very big ones (P.D = 938 (Table 1).
On the day of hatching, the ganglion had a length of 1.200 mm, a volume of 0.2079 mm3 and 18067 cells (P.D = 86902). Among these cells, 4871 (26.96%) were dark type (P.D = 23429) and 13196 (73.04%) were light ones (P.D = 63473). In all, there were 105 (0.58%) tiny cells (P.D = 505), 4179 (D = 428 + L= 3751) (23.13%) were very-small ones (P.D= 20101), 4987 (D = 924 + L = 4063) (27.6%) small ones (P.D= 23987), 6640 (D = 2178 + L= 4462) (36.75%) were medium sized (greatest P.D = 31938), 1531 (D = 693 + L = 838) (8.47%) were big ones (P.D = 7364), 498 (D = 425 + L= 73) (2.76%) were very big ones (P.D = 2395) and 127 (D = 118 + L = 9) (0.7%) were large ones (P.D = 611). In the adult situation, the ganglion had a length of 1.290 mm, a volume of 0.2325 mm3 and 12483 cells (P.D = 53691). Among these cells, 12189 (97.64%) were dark type (P.D = 52426) and 294 (2.36%) were light ones (P.D = 1265). In all, there were 1183 (9.48%) tiny cells (P.D= 5088), 4935 (39.53%) were very small ones (greatest P.D = 21226), 3800 (D = 3645+L = 155) (30.44%) were small ones (P.D= 16344), 1961 (D = 1860 + L= 101) (15.71%) were medium sized ones (P.D = 8434), 182 (D = 168 + L= 14) (1.46%) were big ones (P.D = 783), 295 (D = 278 + L = 17) (2.36%) were very big ones (P.D= 1269) and 127 (D= 120 + L= 7) (1.02%) were large ones (P.D = 546) (Fig. 2).
DISCUSSION
The appearance of a few light cells on Embryonic day 6 (E6) for the first time might indicate an active and early establishment of functional connections around this period.
| Table 1: | Total number of dark and light cells in the vestibular ganglion in different age-groups of animals in the ontogeny of the chick (D = Dark cells, L = Light cells, E = Embryonic age, H = Day of Hatching, A = Adult) |
 | |
This suggestion is based on the following findings. It has been found in the trigeminal ganglion (Hamburger and \Narayanan, 1969) that by the 8th day of incubation at least some of its cells have established peripheral and central connections as indicated by the presence of reflexogenic responses to tactile stimulus of the beak. It is also found thafimany trigeminal sensory cells have extensive peripheral projections by the end of the first week of incubation (Noden, 1980) and that these cells cease dividing very early in embryonic development. Such period coincides with the appearrance of light cells (for the first time) in the trigeminal ganglion during development i.e., on E8 (Pillay, 1999). These light cells have formed as a result of their failure to establish functional connections. It is reasonable to assume that the cells that fail to establish proper central and peripheral connections might become inactive, change to a light-coloured cell on staining. The appearance of light cells on E6 for the first time in the vestibular ganglion during development would explain that the establishment of functional connections begins as early as E6 in this ganglion. This is the earliest stage where the light cells are ever noticed in any one ganglion in the present series of investigation. This in turn emphasise the functional importance of the vestibular ganglion during development and therefore of the vestibular organ. It is quite possible that the embryo and its different organs (especially the vital organs such as heart, lung, digestive system etc) during development must be kept in a suitable position in relation to the gravitational force to insure proper development. That is, the maintenance of proper position of these vital organs in relation to the gravitational force during development should be an important factor for the normal development and growth of the embryo. It is probably because of this reason that the embryo is continuously kept floating in a fluid medium (amniotic fluid) during development in order to allow sufficient freedom to rotate and move around to help maintain a suitable position in relation to gravitational force exerted on the embryo.
There are only a few light-cells (8 cells) on E6. It is interesting to note that the total number of cells continues to increase on E8 (42695 cells) and E10 (61936 cells) (while no light cells are observed) but there is a gross cellular reduction (to 43894 cells) on E13 (while the light cells have made their appearance again). Once again there is an increase in the number of cells on E15 (53439 cells) and E18 (237191 cells) having a rapid proliferation or influx of smaller categories of cells (presymably representing a large number of phagocytes). However, on the day of hatching while the light-cell category has made their appearance for the first time among the very small type of cells, there is again a great reduction or loss in the number of ganglionic-cells.
It is interesting to note from the behavior of the light-cells at different stages of development that the light cells represent a group of resting, inactive, dying, dead or degenerating cells. The occurrence of light cells (for the first time) among the very-small type on the day of hatching suggests that these cells (very-small type) have been actively growing and establishing functional connections till hatching. However, on the day of hatching while the young animal is already prepared for an independent living, there is no need for these very-small cells to grow further because all the functional connections should have been perfectly established by this time. Therefore, it is reasonable to believe that there is no need for these cells to grow further and that they themselves become inactive and turn to be light coloured cells on staining, possibly to disappear from the ganglion.
Other relevant features that could be of interest are discussed below. The results in the vestibular ganglion show that there is a fluctuation in the ganglionic-volume, which does not seem to correspond with the fluctuation in the cell population.
 | |
| Fig. 1: | Dark (D) and Light (L) cells observed in the Ganglion. Different size-categories of cells are also: seen. Each division of the scale-bar is equal to 10 microns |
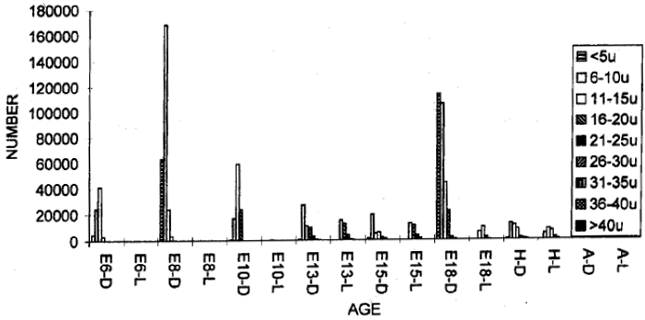 | |
| Fig. 2: | Total number of dark and light cells in the vestibular ganglion in different age groups of animals in the ontogeny of the chick |
The ganglionic-volume on E6 is 0.0094 mm3 that shows gradual increase through E8 and E10 while the cell population is also increasing. The relationship between the size of the cell body, cell number and the volume provides an indication that it is the expansion of the cellular processes that dominates in increasing the size of the ganglion. This suggests that the ganglionic-volume basically increase due to the increase in the size of the neuropil and the size of the cell. It is also evident from earlier investigations (Hamburger and Levi-Montalcini, 1949; Rubel et al., 1976) that the major changes in the cell-size, nuclear size and cell number occur between E13 and the day of hatching in different parts of the central nervous system. The ganglionic-volume increase is minimal during the beginning of development, i.e., up to E10, while as from E13 onwards the volume shows a quicker and greater increase. However, on E13 the cell population has dropped down. There is a coincidence of reduced number of cells (i.e., degeneration) with the appearance of large number of light cells during this period. This is similar to the earlier observation (Rubel et al., 1976) that most cell-death in the brain-stem auditory nuclei occurs in the period between E11 and E13. The ganglionic-volume is gradually increasing except on two occasions: one on E15 while there is a gross reduction in its length in combination with a small decrease in its average cross-sectional diameter and another, on the day of hatching while there is a fractional decrease in its length in combination with a gross reduction in its cross-sectional diameter. However, during the whole embryonic period, the volume reaches its maximum-size (0.3874 mm3) on E18 while the cell population and its density are also the greatest. The ganglionic-volume drops down again to 0.2079 mm3 on the day of hatching but increases only slightly through further growth to reach 0.2325 mm3 in the adult situation. Unlike the conditions observed in many other ganglia (Pillay, 1999), the vestibular ganglion has its greatest volume on E18 and not in the adult situation. However, the rostro-caudal length of the ganglion is greatest only in the adult animal. Such behaviour of the vestibular ganglion having a reduced ganglionic volume in the adulthood could be due to their much-reduced functional capacity due to ageing process as a result of gross reduction in the number of cells.
The size of cell-population on E6 is 13.87 percent that shows an increase through E8, to reach 26.11 percent on E10. This value drops down on E13 to 18.51 percent and increases to 22.53 per cent on E15. Later, there is a sharp increase in the cell-population on E18 with a great influx of smaller categories of cells to gain its greatest value of 100 percent. The first increase in the cell-population observed on E10 concerns mainly with neural elements (nerve cells and neuroglia), whereas the second increase observed on E18 concerns mainly with phagocytic cells (Levi-Montalcini, 1950; Tennyson, 1970) that help remove the unsuccessful neuronal elements. Later these phagocytic cells themselves disappear from the ganglion on the day of hatching. Therefore, the cell population suddenly drops down on the day of hatching while only 7.62 percent cells remain, with a loss of 92.38 per cent cells during this short period. Similar findings of the presence of enormous number of phagocytic cells at corresponding stages have been reported by several investigators (Levi-Montalcini, 1950; Tennyson, 1970; O'Connor and Wyttenbach, 1974; Pilar and Landmesser, 1976). Several investigators (Hamburger and Levi-Montalcini, 1949; Glucksmann, 1951; Saunders, 1966) attributed that the cell death and degeneration could be one of the factors that regulate the size of the cell population and morphogensis during development.
It has been found in the present study that about 45 per cent of cells that have been observed on E6 fail to survive on the day of hatching. This is similar to the earlier reports (Cowan and Wenger, 1967; Rogers and Cowan, 1973; Landmesser and Pilar, 1974; Hamburger, 1975) that 40 per cent or more of the neurones that are initially generated fail to survive to maturity. On further growth of the animal, there is a greater reduction of cells to leave just 5.26 per cent cells in the adulthood that seems to be the lowest value in the whole ontogeny.
ACKNOWLEDGEMENT
The author wishes to express his sincere and unlimited gratitude to Professor. Dr. Dubbeldam A, Leiden University, Netherlands for his continued generosity, guidance, co-operation and support throughout this study. His continued efforts and encouragement to make it as part of a thesis for the award of a DSc degree of Leiden University are greatly appreciated.
REFERENCES
- Cowan, W.M. and E. Wenger, 1967. Cell loss in the trochlear nucleus of the chick during normal development and after radical extirpation of the optic vesicle. Exp. Zool., 164: 267-279.
CrossRefDirect Link - Gaik, G.C. and A.I. Farbman, 1973. The chicken trigeminal ganglion I. An anatomical analysis of the neuron types in the adult. J. Morphol., 141: 43-55.
CrossRefDirect Link - Glucksmann, A., 1951. Cell deaths in normal vertebrate ontogeny. Biol. Rev., 26: 59-86.
CrossRefDirect Link - Hamburger, V. and R. Levi-Montalcini, 1949. Proliferation, differentiation and degeneration in the spinal ganglia of the chick embryo under normal and experimental conditions. J. Exp. Zool., 111: 457-501.
CrossRefDirect Link - Hamburger, V. and C.H. Narayanan, 1969. Effects of the deafferentation of the trigeminal area on the motility of the chick embryo. J. Exp. Zool., 170: 411-426.
CrossRefDirect Link - Hamburger, V., 1975. Cell death in the development of the lateral motor column of the chick embryo. J. Comp. Neurol., 160: 535-546.
CrossRefPubMedDirect Link - Levi-Montalcini, R., 1950. The origin and development of the visceral in the spinal cord of the chick embryo. J. Morphol., 86: 253-283.
CrossRefDirect Link - Landmesser, L. and G. Pilar, 1974. Synaptic transmission and cell death during normal ganglionic development. J. Physiol., 241: 737-749.
Direct Link - Noden, D.M., 1980. Somatotopic and functional organization of the avian trigeminal ganglion: An HRP analysis in the hatchling chick. J. Comp. Neurol., 190: 405-428.
CrossRefDirect Link - O'Connor, T.M. and C.R. Wyttenbach, 1974. Cell death in the embryonic chick spinal cord. J. Cell Biol., 60: 448-459.
CrossRefDirect Link - Pilar, G. and L. Landmesser, 1976. Ultrastructural differences during embryonic cell death in normal and peripherally deprived ciliary ganglia. J. Cell Biol., 68: 339-356.
CrossRefDirect Link - Rubel, E.W., D.J. Smith and L.C. Miller, 1976. Organization and development of brain stem auditory nuclei of the chicken: Ontogeny of N. magnocellularis and N. laminaris. J. Comp. Neurol., 166: 469-489.
CrossRefDirect Link - Rogers, L.A. and M. Cowan, 1973. The development of the mesencephalic nucleus of the trigeminal nerve in the chick. J. Comp. Neurol., 147: 291-319.
CrossRefDirect Link - Spassova, I., 1982. Cat trigeminal ganglion. Neuron types. An experimental study. Zeitschrift Mikroskopisch-Anatomische Forschung, 96: 235-244.
PubMed - Tennyson, V.M., 1970. The fine structure of the axon and growth cone of the dorsal root neuroblast of the rabbit embryo. J. Cell Biol., 44: 62-79.
CrossRefPubMedDirect Link