
Amal A. H. Saleh
Botany Department, Faculty of Science, Suez Canal University, Ismailia, Egypt
Pakistan Journal of Biological Sciences
Year: 2002 | Volume: 5 | Issue: 4 | Page No.: 455-460







ABSTRACT
Chorcorus olitorius grows in nutrient solution was treated with Ni (10 and 50μM) and EDTA (10, 50 and 100 μM) in different combinations. Shoot, root length, fresh and dry weight for shoot and root photosynthetic pigments and carbohydrates were decreased with increasing Ni supply. EDTA reduces the inhibition of Ni on chlorophyll and total carbohydrates formation. Proline content was increased significantly with increasing Ni concentration and decreased after EDTA treatment. Ni and EDTA treatment induced quantitative and qualitative changes in the protein PAGE profile of plant. New polypeptides of varying molecular weights were synthesized with the addition of Ni and others were stimulated by Ni and EDTA to nutrient solution. In general, the total number of protein bands were decreased by Ni treatment but increased after EDTA addition. It was noticed that, increasing Ni concentration in nutrient solution significantly increased uptake and accumulation of Ni in both shoot and root.The concentration of Ni in shoot and root was greater than in shoot. Accumulation of K, Ca and Fe were significantly decreased with increasing Ni concentration. The uptake of Ni is diminished in the presence of EDTA, suggesting that the Ni-EDTA complex is unable to penetrate the membrane. Increasing EDTA concentration combined with Ni, increased the concentration of Ca, K and Fe. In conclusion EDTA can ameliorate the toxic effects of Ni on Chorcorus olitorius plant.
PDF Abstract XML References Citation
How to cite this article
Amal A. H. Saleh, 2002. Response of Anabolic Capacities, Proline, Protein Patterns and Mineral Elements to Nickel and EDTA Stress in Chorcorus olitorius. Pakistan Journal of Biological Sciences, 5: 455-460.
DOI: 10.3923/pjbs.2002.455.460
URL: https://scialert.net/abstract/?doi=pjbs.2002.455.460
DOI: 10.3923/pjbs.2002.455.460
URL: https://scialert.net/abstract/?doi=pjbs.2002.455.460
INTRODUCTION
Soil contamination with heavy metals has become a worldwide problem, leading to a loss in agricultural yield and hazardous health effects as they enter the food chain (Nellessen and Fletghe, 1993; Guo and Marschner, 1995). Man’s energy and chemical consumption is the main cause of trace element pollution in the biosphere. The main sources of contamination in agricultural soils are fertilizer impurities and the use of refuge-derived compost and sewage sludge (Ni2+). Disposal of household, municipal and industrial waste has given rise to even more widespread nickel soil concentrations (Alloway, 1995). Nickel has recently been defined as essential micronutrient, because of its involvement in enzymatic activity in legumes (Welch and Shuman, 1995). In most plants it is found at the level of 0.1-5 ppm (on a dry weight basis), with a wide threshold range of 40-246 ppm for toxicity symptoms, dependent upon the plant species (Kabata-Pendias and Pendias, 1992). The main common symptoms are chlorosis, and inhibited photosynthesis and respiration (Bazzaz et al., 1974, Foy et al., 1978). The mechanisms governing Ni2+ toxicity are not well understood. The dry matter of wheat was not affected by Ni treatment (Zeller and Feller, 1999). Nickel prevented any growth of redbeet plant (Kukier and Chaney, 2001). Cd caused reduction in dry weight and carbohydrate concentration in sugar beet plant (Greger and Bertell, 1992). EDTA reduces the inhibition of Cd2+ on sugar formation and accumulation (Greger and Lindberg, 1987). Exposure of Indian mustard to high concentrations of Pb and EDTA caused reductions in both the transpiration rate and the shoot water content (Andrew et al., 1998). Excess EDTA to nutrient cations, reduce the root damage (Eskew et al., 1984). After EDTA treatment, soluble heavy metals in soil water occurred mainly as metal-EDTA complexes and high concentrations of heavy metals in soil water after EDTA treatment could pose an environmental risk in the form of ground water contamination (Lambi et al., 2001). Many plant species accumulate high concentrations of proline under stress conditions such as salt, drought, or heavy metals stress. Furthermore, proline serves as a compatible solute to protect macromolecular structures, as a radical scavenger, or as a rapid source of energy for recovery from stress (Hare and Cress, 1997; Nanjo et al., 1999; Chakravarty and Srivastava, 1997). The concentration of free proline increased in sunflower and nontolerant ecotype of Silene vulgaris leaves in the presence of heavy metals (Kastori et al., 1992). The function of proline under stress conditions is not fully understood. The mobility of Ni2+ is pH dependent (Adersson and Nilsson, 1974; Baker et al., 1979) and other cations and chelating agents (EDTA) can reduce the mobility (Hardiman and Jacoby, 1984). Heavy metals decreased protein content and delay in the synthesis of certain polypeptides (Krupa, 1988 and Jana et al., 1987). Increase in soil solutions concentrations of Ni resulted in increased Ni concentrations in plant tissues (Patrick et al., 1989; Zeller and Feller, 1999; Brown et al., 1989) and the accumulation of heavy metals was more intensive in root than in the shoot of sunflower (Kastori et al., 1992). High heavy metals concentration reduced the iron (Fe) content of the tissues and total uptake of Fe in Sorghum (Kue and Mikkelsen, 1981). EDTA in hydroponics solutions caused significant inhibition of Cd accumulation by the roots and increased Fe concentrations in the shoots (Checkai et al., 1987 and Chaney et al., 1972). Ni+2 contamination in cucumber plants hinders the transport of K and Zn and leads to a higher accumulation of Mn in the roots (Varga et al., 1999). The purpose of this study was to investigate the effect of different Ni2+ concentrations and EDTA on the growth, some metabolic products, electrophoratic patterns of proteins, nickel and some nutrient elements uptake by Chorcorus olitorius plant. Also, tried to eleviate the symptoms of Ni2+ toxicity by using different concentrations of EDTA.
MATERIALS AND METHODS
Plant culture: Seeds of Chorcorus olitorius were obtained from the Field Crop Institute, Agricultural Research Center, Egypt. Seeds were germinated in pots containing acid washed sandy-loam soil for two weeks. After germination, the seedlings were transferred to pots containing nutrient solution based on half strength Hoagland solution at pH 6.8 for ten days of preculture. The nutrient solutions were constantly aerated and replaced every 2 days. The seedlings were kept in Precision low temperature incubator Model 818. 25/27 oC night/day temperature, 63-65 % relative humidity under 12 hrs photoperiod and the light intensity was 1100 Lux. This search was carried out in Botany Department, Faculty of Science, Suez Canal University during July and August 2001.
Exposure to Ni2+: Seedlings were transferred to pots with fresh nutrient solution containing different concentrations of Ni (0, 10 and 50 μM). Ni was added as NicL2.6H2O. The nutrient solution were constantly aerated and replaced every 2 days.
Exposure to Ni2+ and EDTA: In a second experiment, to assess the effect of Ni and EDTA (ethylene diamine tetraacetic acid) on toxicity, seedlings were transferred to pots contained nutrient and different combinations of Ni (0, 10 and 50 μM) and EDTA (0,10,50 and 100 μM). EDTA was added as dipotassium salt. Plants were harvested separately after 2 weeks treatment.
Growth parameters: Four plants from each treatment were chosen randomly for the measurement of root and shoot length, fresh and dry weights for root and shoot. The plants were dried to constant weight at 70 °C to estimate the total carbohydrates and mineral elements content (K, Ca, Fe and Ni).
Determination of photosynthetic pigments: The concentration of pigments as μg/g was determined spectrophotometrically in fresh leaves according to Metzner et al. (1965).
Determination of total carbohydrates: Total carbohydrate contents were extracted according to Dubois et al. (1956) and estimated colorimetrically by the phenol-sulphoric acid method. The results were expressed as mg glucose/100mg dry weight.
Determination of proline: The proline content as μ mols/100 gm plant tissue was extracted with aqueous sulphosalicylic acid and estimated spectrophotometrically in fresh shoot system.
Protein electrophoresis
Extraction of total proteins: Total protein extracts were prepared by extracting appropriate weight from the frozen plant material with 0.125 M tris/borate, pH 8.9. All the obtained extracts were kept at 4°C for 24 hr and then centrifuged at 10,000 rpm for 20 min. The supernatants were used for electrophoresis.
Gel electrophoresis: SDS polyacrylamide gel electrophoresis (PAGE) was carried out with gel slabs according to the method of Laemmli (1970). Protein subunit bands were stained with coomassie blue R-250 by standard techniques. The gel was scanned using Jel-Pro-Analyzer ver. 3.3 (Media Cybermetics, 93-97).
Determination of minerals in plant material: The wet ashing method described by Westerman (1990) was used for the estimation of Ni, Ca, K and Fe concentrations. Analysis was carried out using a Perkin-Elmer 3100 atomic absorption spectrophotometer. The data were recorded in ppm dry weight.
Statistical analysis: All parameters were statistically analyzed by a one way and two ways ANOVA followed by least significant difference (L.S.D.) at 5% level (p=0.05). Also, Pearson Rank Correlation test was carried out at SPSS version 8. (R=correlation or regression value).
RESULTS
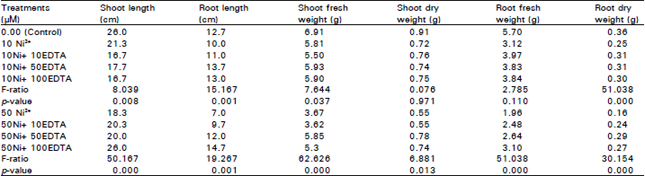
Shoot and Root length, fresh and dry weight: When the seedlings of Chorcorus olitorius were treated with Ni2+, shoot and root length, fresh and dry weight decreased with increasing Ni2+ concentration in comparison with the control (Table 1). With the addition of EDTA to nutrient solution containing 50 μM Ni2+, all the above growth parameters increased significantly with increasing EDTA concentration. At 10 μM Ni2+, root length correlated increased significant with increasing EDTA concentration (R=0.736, P=0.006) and other growth parameters increased but not significantly.
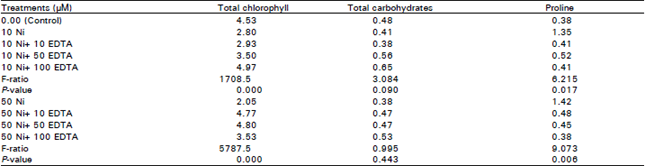
Photosynthetic pigments: The results in Table 2 show that, the total chlorophyll decreased significantly at 10 and 50 μM Ni2+ by 38.1 and 54.8 % respectively in comparison with the control. The chlorophyll content correlated significantly and increased with increasing EDTA concentration combined with 10 and 50 μM Ni2+ (R=0.982, 0.192 & P=0.000, 0.547 respectively).
Total carbohydrates: In Chorcorus olitorius plants grown in nutrient solution containing 10 μM Ni2+, the carbohydrate content decreased but not significant and significantly decreased at 50 μM Ni2+ by 20.8% compared with the control. An increase in the supply of EDTA combined with 10 and 50 μM Ni2+, the carbohydrate concentration significantly increased gradually (Table 2) (R=0.934, 0.735& P=0.000, 0.001 respectively).
Proline content: Table 2 shows that the proline content in Chorcorus olitorius significantly increased with increasing Ni concentration compared with the control. In the presence of EDTA in nutrient solution containing 10 and 50 μM Ni2+, the proline content correlated and significantly decreased (R=-0.586, -0.647& P=0.016, 0.006 respectively).
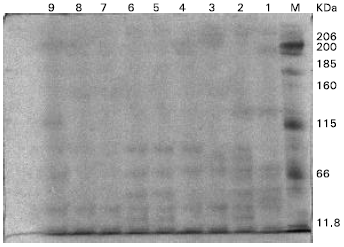 | |
| Fig. 1: | Electrophotograph of Sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) of the total proteins of Corchorus olitorius plants, 1= untreated plants (control) (track1); 2= plants treated with 10 μM Ni2+ (track 2); 3,4 &5= plants treated with 10,50 and 100 μM EDTA combined with 10 μMNi2+ (tracks 3,4, & 5 respectively); 6= plants treated with 50 μM Ni2+ (track 6); 7,8 &9= plants treated with 10, 50 & 100 μM EDTA combined with 50 μM Ni2+ (tracks 7,8 and 9 respectively); M= Marker. |
Protein electrophoresis: The gel electrophoretic technique revealed both quantitative (band intensity and relative mobility) and qualitative (disappearance of some bands or appearance of new bands) changes in SDS-PSGE protein patterns of Chorcorus olitorius plant in response to Ni2+ and nickel combined with different concentrations of EDTA (Table 3 and Fig. 1). In general, the number of protein bands decreased at 10 and 50 μM ( 4 and 5 bands respectively). These bands increased when the plants treated with EDTA Ni2+ in combination. It was noticed that, the band number increased at 50 μM Ni2+ combined with 10, 50 and 100 μM EDTA (8,11 and 10 bands respectively). But when the plants were treated with 10 μM Ni2+ combined with 50 and 100 μM EDTA, the total bands of protein were 7 , 10 and 50 μM Ni2+ completely inhibited the synthesis of a 129.4 KDa polypeptide, but EDTA (50 and 100 μM) combined with 50 μM Ni2+ were able to synthesizes this polypeptide again (149.40 KDa). With addition of 50 and 100 μM EDTA to nutrient solution containing 10 μM Ni2+, the new polypeptides were synthesized at 90.46, 72.4 and 16.46 KDa and the concentration of these polypeptides were increased with increasing EDTA concentration. The highest Ni2+ concentration (50μM) added to nutrient solution containing EDTA induced the synthesis of new polypeptides at 236.65, 227.68, 216.57, 202.75, 129.40, 83.36 and 66.0 Kda compared with lower Ni concentration (10μM).
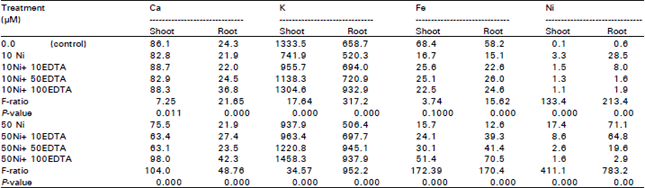
Minerals in plant tissues: Table 4 shows that, increasing Ni2+ concentration in nutrient solution strongly increased the concentration of Ni2+ in both cshoot and root of Chorcorus olitorius. The concentration of Ni was greater in roots than shoots and was statistically significant. Ca, K and Fe concentrations were decreased in shoot and root of Chorcorus olitorius with increasing Ni concentration in nutrient solution. It was noticed that, the concentrations of Ca and K were greater in shoots than roots.
| Table 1: | Effect of different concentrations of Ni2+ (μΜ) and EDTA (μΜ) in nutrient solution on vegetative growth parameters of Chorcorus olitorius plant. Values are means of three replica |
 | |
| Table 2: | Effect of different concentrations of Ni2+ (μΜ) and EDTA (μΜ) in nutrient solution on total chlorophyll (mg/g fresh wt.), total carbohydrates (mg glucose/100mg dry wt.) and proline content (μM/100 g fresh wt.) of Chorcorus olitorius plant |
 | |
| Table 3: | Comparitive analysis of relative concentrations, molecular weight and mobility rate (Rm) of proteins in Chorcorus olitorius plants treated with different combinations of Ni2+ and EDTA.These bands were separated using SDS-PAGA technique. |
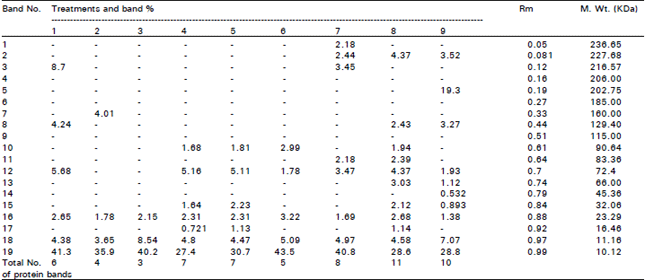 | |
Treatments: 1=untreated plants (control); 2=10μM Ni2+; 3,4 &5= 10, 50 &100 μM EDTA+10μM Ni2+ respectively; 6= 50μM Ni2+; 7,8&9=10, 50 &100 μM EDTA+50μM Ni2+ respectively; | |
Increasing the concentration of EDTA combined to Ni increased the concentration of Ca, K and Fe in both shoots and root and decreased Ni concentrations in Chorcorus olitorius tissues.
DISCUSSION
The present data (Table 1) showed that, shoot and root length, fresh and dry weight decreased at 10 and 50μM Ni2+ due to the inhibitory effect of heavy metals on cell division and cell elongation and enzyme activity (Balsberg, 1989). Nickel prevented any growth of redbeet plant (Kukier and Chaney, 2001). Also, this decline in growth due to an accumulation of heavy metals in plant tissues (Bonnet et al., 2000). With the addition of EDTA to nutrient solution containing Ni2+, the growth parameters increased with increasing EDTA concentration. Chelators may protect plants from damage caused by heavy metals. EDTA reduced Ni2+ toxicity (Singh and Pandy, 1981).
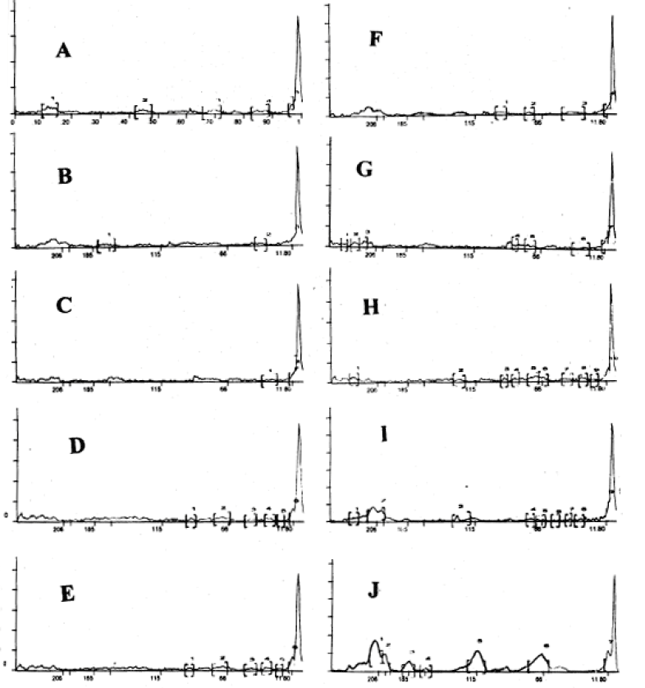 | |
| Fig. 2: | Scans of the tracks in Fig. 1 A = scan of the track M (mol. Wt. Markers); B= scan of the track 1 (control); C= scan of the track 2 (10μM Ni2+); D, E, & F = scan of the tracks 3, 4, & 5 (10, 50 & 100 μM EDTA combined with 10 μM Ni2+); G= scan of the track 6 (50μM Ni2+); H, I, & J= scan of the tracks 7, 8 & 9 (10, 50 & 100 μM EDTA combined with 50 μM Ni2+). |
| Table 4: | Effect of different concentrations of Ni2+ (μM) and EDTA (μM) in nutrient solution on Ca, K and Fe content in plant tissues of Chorcorus olitorius plant. Values expressed as ppm/dry weight |
 | |
Excess EDTA to nutrient solution, reduce the root damage (Eskew et al., 1984).
The results of photosynthetic pigments in Table 2 showed that, chlorophyll content decreased with increasing Ni concentration in nutrient solution. The main common symptoms of heavy metals are chlorosis and inhibited photosynthesis and respiration (Bazzaz et al., 1974 and Foy et al., 1978). In Brassica nepus, heavy metals significantly decreased root dry weights and totals chlorophyll contents (Larsson et al., 1998). In wheat, the inhibited growth by increased heavy metal concentrations was attributed to indirect effects of heavy metals on contents of essential nutrients or structural damages of chloroplasts (Ouzounidou et al., 1997). Chlorosis occurs due to Fe and Mg deficiency symptoms (Greger and Lindberg, 1987). Mg is necessary for chlorophyll formation and Fe is needed in ferredoxins in the photosynthetic redox system (Khan and Khan 1983). Also, the chlorophyll synthesizing system and chlorophyllase activity were affected by heavy metals (Jana et al., 1987). EDTA increased the chlorophyll content when added to nutrient solution containing Ni2+ due to the formation of heavy metal-EDTA complex and this complex is unable to penetrate the plant membrane (Greger and Lindberg, 1987). Also, EDTA reduces the mobility of heavy metal and then decreases its toxicity (Hardiman and Jacoby, 1984). The present investigation shows that, carbohydrate formation is inhibited by Ni (Table 2).This could be an effect of a reduced photosynthesis as the contents of all the investigated sugars decrease (Vallee and Ulmer, 1972). Cd caused reduction in dry weight and carbohydrate concentration in sugar beet plant (Greger and Bertell, 1992). This could be an effect of a reduced photosynthesis as the contents of all the investigated sugars decrease (Kremer and Markham, 1982). EDTA reduces heavy metal toxicity (Singh and Pandy, 1981) which agrees with this experiment that the Ni inhibition on the carbohydrate content in Chorcorus olitorius was suppressed by EDTA (Table 2) due to the formation of heavy metal-EDTA complex and this complex is unable to penetrate plant membrane (Greger and Lindberg, 1987).
In Chorcorus olitorius treated with Ni2+, the proline content was increased. The concentration of free proline increased in sunflower leaves and Silene vulgaris in the presence of heavy metals (Kastori et al., 1992 and Kastori et al., 1992). The possibility of proline involvement in tolerance mechanisms to heavy metals (Raman and Shanti, 1999). Sharp rise in free proline under stress due to protein proteolysis and the activities of protein biosynthetic enzymes (Chang and Lee, 1999). EDTA reduces the proline concentration due to the formation of heavy metal-EDTA complex.
The present study shows that, the number of protein bands decreased when the plant was treated with Ni2+. Heavy metals caused a delay in synthesis of certain polypeptides (Krupa, 1988). The decrease in protein content revealed that soluble proteins may be leaked or diffused out of the plant material, or possibly the catabolic enzymes were induced and destroyed the protein (Jana et al., 1987). These bands increased by addition of EDTA to nutrient solution containing Ni. The plants synthesize new polypeptides to alleviate Ni-stress effect or detoxification of Ni. The synthesis of certain polypeptide of specific molecular weights has been known as metal-binding polypeptides (Jackson et al., 1985; Rauser, 1984; Reese and Wgner, 1987; El-Enany and Abd-Allah, 1995).
The content of Ni increased in both shoot and root of Chorcorus olitorius as the Ni supply increased (Table 4) and its concentration in root was more than in shoot. Increase in soil solutions concentrations of Ni resulted in increased Ni concentrations in plant tissues (Patrick et al., 1989; Zeller and Feller, 1999; Brown et al., 1989) and the accumulation of heavy metals was more intensive in root than in shoot of sunflower (Kastori et al., 1992). Concentration of Ca, K and Fe was decreased in shoot and root of Chorcorus olitorius with increasing Ni concentration in nutrient solution). Heavy metals inhibited the absorption and accumulation of K, Ca and Fe in plant tissues (Trvedi and Erdei, 1991; Woolhouse, 1983; Boardman and McGuire, 1990 and Simon et al., 1994). Zhang et al. (1991) and Nicolaus et al. (1999) showed that mobilization of Fe by phytosiderophores slightly inhibited by Zn and strongly inhibited by Cu. Nicotinamine (NA) occurs in all plants and chelates metal cations include Fe+2 and have an important role in scavenging Fe and protecting cell from oxidative damage. This reduction of Fe in shoot resulting from coating occurred on root surface and intensified with increasing heavy metal concentrations in the substrate or could be attributed to the formation of high valent heavy metal oxides on the root surfaces which may certain Fe and reduce its absorption by sorghum. The presence of EDTA in nutrient solution diminished the Ni uptake probably by chelating Ni. Hardiman and Jacoby (1984) obtained similar results on bush beans. The uptake of heavy metals is reduced in the presence of EDTA, suggesting that heavy metal-EDTA complex formation is unable to penetrate the membrane (Greger and Lindberg, 1987). In nutrient solution containing FeEDTA, other metals may form their respective EDTA chelates by displacing some Fe from FeEDTA. These reactions may lower the concentration of the free cations of many micronutrient metals while increasing the concentration of Fe (Eskew et al., 1984).
From the results of this experiment, it could be concluded that, Chorcorus olitorius have the ability to uptake and accumulate Ni from the nutrient solution in its tissues to toxic level and DETA ameliorate this toxic effect of Ni.
REFERENCES
- Adersson, A. and K.O. Nilsson, 1974. Influence of lime and pH on Cd availability to plants. Ambio, 3: 198-200.
Direct Link - Andrew, D., K. Yoram, R. Iiya and E. David, 1998. The role of EDTA in lead transport and accumulation by Indian Mustard. Plant Physiol., 117: 447-453.
PubMedDirect Link - Baker, D.E., M.C. Amacher and R.M. Leach, 1979. Sewage sludge as a source of cadmium in soil-plant-animal systems. Environ. Health Perspect, 28: 45-49.
PubMedDirect Link - Pahlsson, A.M.B., 1989. Toxicity of heavy metals (Zn, Cu, Cd, Pb) to vascular plants: A literature review. Water Air Soil Pollut., 47: 287-319.
CrossRefDirect Link - Chakravarty B., S. Srivastava, 1997. Effect of genotype and explant during in vitro response to cadmium stress and variation in protein and proline content in linseed. Ann. Bot., 79: 487-491.
Direct Link - Bonnet, M., O. Camares and P. Veisseire, 2000. Effects of zinc and influence of Acremonium lolii on growth parameters, chlorophyll a fluorescence and antioxidant enzyme activities of ryegrass (Lolium perenne L. cv Apollo). J. Exp. Bot., 51: 945-953.
CrossRefDirect Link - Brown, P., L. Dunemann, R. Shulz and H. Marschner, 1989. Influence of redox potential and plant species on the uptake of nickel and cadmium from soils. Z. Pflanzenernahr Bodenk, 152: 85-91.
CrossRefDirect Link - Chaney, R.L., J.C. Brown and L.O. Tffin, 1972. Obligatory reduction of ferric chelates in iron uptake of soybean. Plant Physiol., 50: 208-213.
PubMedDirect Link - Chang, Y. and T. Lee, 1999. High temperature-induced free proline accumulation in Gracilaria tenuistipitata (Rhodophyta). Bot. Bull. Acad. Sin., 40: 289-294.
Direct Link - Checkai, R., R. Corey and P. Helmke, 1987. Effects of ionic and complexed metal concentrations on plant uptake of cadmium and micronutrient metal from solution. Plant Soil, 99: 335-345.
CrossRefDirect Link - Foy, C.D., R.L. Chaney and M.C. White, 1978. The physiology of metal toxicity in plants. Ann. Rev. Plant Physiol., 29: 511-566.
CrossRefDirect Link - Greger, M. and G. Bertell, 1992. Effect of Ca and Cd on the carbohydrate metabolism in sugar beet (Beta vulgaris). J. Exp. Bot., 43: 167-173.
Direct Link - Guo, Y. and H. Marschner, 1995. Uptake, distribution and binding of cadmium and nickel in different plant species. J. Plant Nutr., 18: 2691-2706.
CrossRefDirect Link - Hardiman, R.T. and B. Jacoby, 1984. Absorption and translocation of Cd in bush beans (Phaseolus vulgaris L.). Physiol. Plant., 61: 670-674.
CrossRefDirect Link - Hare, P.D. and W.A. Cress, 1997. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul., 21: 79-102.
CrossRefDirect Link - Kastori, R., M. Petrovic and N. Petrovic, 1992. Effect of excess lead, cadmium, copper and zinc on water relations in sunflower. J. Plant Nutr., 15: 2427-2439.
CrossRefDirect Link - Khan, S. and N.N. Khan, 1983. Influence of lead and cadmium on the growth and nutrient concentration of tomato (Lycopersicum esculentum) and egg-plant (Solanum melongena). Plant Soil, 74: 387-394.
CrossRefDirect Link - Kue, S. and D. Mikkelsen, 1981. Effect of P and Mn on growth response and uptake of Fe, Mn and P by Sorghum. Plant Soil, 62: 15-22.
CrossRefDirect Link - Kukier, U. and R. Chaney, 2001. Amelioration of nickel phytotoxicity in Muck and mineral soils. J. Environ. Qual., 30: 1949-1960.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lambi, E., F.J. Zhao, S.J. Dunham and S.P. McGrath, 2001. Phytoremediation of heavy metal-contaminated soils. J. Environ. Qual., 30: 1919-1926.
CrossRefDirect Link - Nanjo, T., M. Kobayashi, Y. Yoshiba, Y. Sanada and K. Wada et al., 1999. Biological functions of proline in morphogenesis and osmotolerance revealed in antisense transgenic Arabidopsis thaliana. Plant J., 18: 185-193.
PubMed - Nellessen, J.E. and J.S. Fletcher, 1993. Assessment of published literature pertaining to the uptake/accumulation, translocation, adhesion and biotransformation of organic chemicals by vascular plants. Environ. Toxicol. Chem., 12: 2045-2052.
CrossRefDirect Link - Von Wiren, N., S. Klair, S. Bansal, J.F. Briat and H. Khodr et al., 1999. Nicotianamine chelates both FeIII and FeII. Implications for metal transport in plants. Plant. Physiol., 119: 1107-1114.
CrossRefDirect Link - Ouzounidou, G., M. Moustakas and E.P. Eleftheriou, 1997. Physiological and ultrastructural effects of cadmium on wheat (Triticum aestivum L.) leaves. Arch. Environ. Contam. Toxicol., 32: 154-160.
CrossRef - Raman, B. and S. Shanti, 1999. Changes in proline content accompanying the uptake of zinc and copper by Lemna minor. Ann. Bot., 72: 151-154.
Direct Link - Rauser, W.E., 1984. Isolation and partial purification of cadmium-binding protein from roots of grass Agrostis gigantia. Plant Physiol., 74: 1025-1029.
Direct Link - Reese, R.N. and G.J. Wagner, 1987. Properties of tobbaco Nicotiana tabacum cadmium-binding peptide(s). Unique non-metallothionein cadmium ligands. Biochem. J., 241: 641-647.
Direct Link - Jana, S., T. Dalal and B. Barua, 1987. Effects and relative toxicity of heavy metals on Cascuta reflexa. Water Air Soil Pollut., 33: 23-27.
CrossRefDirect Link - Simon, L., T. Smalley and J. Jones, 1994. Aluminum toxicity in tomato: 1. Growth and mineral nutrition. J. Plant Nutr., 17: 293-306.
Direct Link - Singh, S.P. and A.K. Pandey, 1981. Cadmium toxicity in a cyanobacterium: Effect of modifying factors. Environ. Exp. Bot., 21: 257-265.
Direct Link - Trvedi, S. and L. Erdei, 1991. Effect of cadmium and lead on the accumulation of Ca2+ and K+ and on the influx and translocation of K+ in wheat of low and high K+ status. Physiol. Plant., 84: 94-100.
CrossRefDirect Link - Vallee, B.L. and D.D. Ulmer, 1972. Biochemical effects of mercury, cadmium and lead. Annu. Rev. Biochem., 41: 91-128.
CrossRefPubMedDirect Link - Welch, R.M. and L. Shuman, 1995. Micronutrient nutrition of plants. Crit. Rev. Plant Sci., 14: 49-82.
CrossRefDirect Link - Zeller, S. and U. Feller, 1999. Long-distance transport of cobalt and nickel in maturing wheat. Eur. J. Agron., 10: 91-98.
Direct Link - Zhang, F., M. Treeby and H. Marschner, 1991. Mobilization of iron by phytosiderophores as affected by other micronutrient. Plants Soil, 130: 173-178.
CrossRefDirect Link - Larsson, E., J. Borman and H. Asp, 1998. Influence of UV radiation and Cd2+ on chlorophyll florescence, growth and nutrient content in Brassica napus. J. Exp. Bot., 49: 1031-1039.
CrossRef - Greger, M. and S. Lindberg, 1987. Effects of Cd2+ and EDTA on young sugar beets Beta vulgaris. II- Net uptake and distribution of Mg2+, Ca2+ and Fe2+/ Fe2+. Physiol. Plant, 69: 81-86.
CrossRefDirect Link - Varga, A., R.M.G. Martinez, G. Zaray and F. Fodor, 1999. Investigation of effects of cadmium, lead, nickel and vanadium contamination on the uptake and transport processes in cucumber plants by TXRF spectrometry. Spectrochimica Acta Part B: Atomic Spectroscopy, 54: 1455-1462.
CrossRefDirect Link