
A. M. Gaber
Botany Department, Faculty of Science, Mansoura University, Mansoura, Egypt
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 5 | Page No.: 514-524







ABSTRACT
The effect of water stress induced by salinity (0.25 and 0.5 % NaCl) or excessive water supply (wet and waterlogged treatments) on growth, photosynthetic activity, carbohydrates, nitrogen, mineral elements (Na, K, Ca, Mg, Mn, Fe) and some organic acids (Oxalic and citric) as well as keto acids (pyruvic and -ketoglutaric acid) were studied. A significant increase in growth criteria (shoot length , shoot fresh and dry weight, total green leaf area, specific leaf area, net assimilation rate and relative growth rate of Vigna sinensis shoot was detected in response to 0.25% NaCl and wet treatment above the respective controls, while the highest level of salinity (0.5%) and waterlogging seemed to have a decreasing effect in the previous growth parameters. Salinity stress markedly decreased the soluble sugars but greatly increased polysaccharides and total carbohydrates. On the other hand, different water treatments (wet and waterlogging) reduced significantly the carbohydrate fractions (soluble sugars, sucrose, polysaccharides and total carbohydrates). 14CO2 photoassimilation indicated that soluble, insoluble and consequently total photosynthates were reduced under the effect of water stress. A significant decrease in total-N and protein-N was observed in relation to control by higher level of salinity (0.5%) and waterlogging whereas the soluble-N was, in general, increased significantly. Meanwhile, moderate stress induced by 0.25% NaCl and wet treatment showed a non-significant responses in nitrogen fractions. Mineral contents of the tested plant (Vigna sinensis) were affected greatly by water stress. The levels of K, Ca, Mg, Mn and Fe were reduced greatly whereas Na showed a reversible situation. Waterstress induced by salinity resulted in a pronounced increase in pyruvic acid whereas the -ketoglutaric acid showed a reversible situation compared to control. Wet and waterlogging treatments decreased these two keto acids below the respective controls. Oxalic and citric acids content of Vigna sinensis shoot were decreased significantly by water stress as induced by salinity or different water treatments. Foliar application of IAA (100 ppm) to the Vigna sinensis plant at the vegetative stage or presoaking the seeds in oxalic acid (100 ppm) equilibrated, if any, the suppressive effect of water stress on growth and the metabolic activities in shoot of water stressed plant. The result revealed that IAA and/or oxalic acid alleviate the stress symptoms and regulate the changes in growth and some metabolic activities of salinity and waterlogging.
PDF Abstract XML References Citation
How to cite this article
A. M. Gaber, 2003. Response of Water Stressed Vigna sinensis Plants to Application of IAA and/or Oxalic Acid. Pakistan Journal of Biological Sciences, 6: 514-524.
DOI: 10.3923/pjbs.2003.514.524
URL: https://scialert.net/abstract/?doi=pjbs.2003.514.524
DOI: 10.3923/pjbs.2003.514.524
URL: https://scialert.net/abstract/?doi=pjbs.2003.514.524
INTRODUCTION
Stress conditions can rapidly and dramatically interfere with plant growth and metabolism. Waterlogged soils adversely affect the growth of plants not adapted to wet land conditions, also salinity can affect growth and water content of several plants species (Younis et al., 1999; Mamdouh et al., 2002).
Plant growth is often suppressed as a result of root zone stress such as salinity, drought and anaerobic conditions (Carmi, 1993). Water stress induced by water drought or waterlogging not only shows several physiological and biochemical disturbances associated with general reduction in size but also exhibits characteristic modifications in structure; particularly of leaves. The symptoms in waterlogging are similar to those reported for drought stress although the mechanism of causes may be different (Pelleschi et al., 1997; Younis et al., 1999).
Photosynthesis is an important process, which can be altered by waterlogging or salinity stress. A decrease in CO2 assimilation was induced by waterlogging (Younis et al., 1993; Habib-ur-Rehman, 1999) and salinity (Sultana et al., 1999).
Water stress can lead to change in carbohydrates production. Carbon metabolism may have some contribution to salinity tolerance in plant (Rout and Shaw, 1998). Decrease in glucose, fructose, polysaccharides and total carbohydrates were induced in Vicia faba shoots by waterlogging (El-Shahaby et al., 1994) and Phaseolus vulgaris in shoots and roots by salinity (Younis et al., 1993). On the other hand, Imamul-Hauq and Larher (1983) found that salinity increased total soluble sugars in Phaseolus aureus shoots.
Agarwal and Gupta (1995) showed that water stress may be alleviated by using certain bioregulators such as IAA which suppressed the water stress and in turn its injuries by lowering osmotic potential and maintaining cellular turgidity. In the meantime, proline, soluble amino acids, soluble sugar, insoluble sugar and proteins were increased by this growth regulators. Moreover, Pustovoitova et al. (2000) stated the higher water stress tolerance of the transgenic tobacco plants (Nicotiana silverstris) is related to IAA involvement in plant adaptations, i.e. in the hormone induced modification of plant hormonal status and the osmoregulation process. Plant hormones have much to offer in improving crop qualitatively and quantitatively. In the meantime, plant hormones are a useful tool in saving irrigation (Yang and Clore, 1973).
Organic acids have been implicated in various roles in the metabolic and physiological responses of plants to water stress (Franco et al., 1992).
The role of organic acids in water stress is well investigated by Timpa et al. (1986) and Marschner (1995), they reported that the synthesis and breakdown of organic acids serve as a mechanism for pH regulation in plant cell.
Hamdia et al. (1996) stated that the presoaked seeds of onion and maize with some organic acids (oxalic, citric and/or succinic) counteracted the suppressive effect of the relatively higher salinity levels.
The objective of this study was to investigate the possibility of using IAA and/or oxalic acid, either singly or incombination, for improvement of growth and metabolic activities of water stressed Vigna sinensis plant.
MATERIALS AND METHODS
Plant materials and growth conditions: Pure strains of Vigna sinensis (Southern pea var. Cream 7) were obtained from the Agricultural Research Center, Ministry of Agriculture, Egypt. The seeds after surface sterilization, were thoroughly washed and then soaked in distilled water. Thereafter, 12 seeds of Vigna sinensis were sown separately in plastic pots (25 cm in diameter) containing equal amounts (4.5 kg) of homogeneous mixture of sand and clay soil (1:2 w/w). The seedling were then left to grow in a green house under normal day/night conditions. Uniform watering was carried out. Thinning was carried out after 14 days to leave 5 uniform plants of Vigna sinensis per pot.
The pots were divided into 6 groups
Control: In control pots, the water holding capacity was kept at 60% of the total water holding capacity.
Wet and waterlogging treatments: In this group, the water levels were 100 and 140% of water field capacity of soil. Water holding capacity of the used mixture of soil determined as g (%) of dry soil was 30%. To maintain the field capacity characteristic of each of the two different water treatments (wet and waterlogging), watering was always made when the field capacities were lowered to 30 and 42% for wet and waterlogged soil, respectively.
Saline treatments: The pots of this group were aerated and treated with NaCl at two levels (0.25% and 0.5%). For maintaining the salinity at each desired levels, the soil in each pot was initially irrigated by saline solution (NaCl) so that the required percent of salinity was obtained. Thereafter, the pots were irrigated occasionally with certain amounts of tap water in order (1) to keep the water content at 60% of the total water holding capacity of the soil used and (2) to allow the level of soil salinity in each pot to be unchanged.
IAA treatment: Each previous treatment was performed with the simultaneous foliar spray of 100 ppm IAA, each pot took 100 ml.
Oxalic acid treatment: In this group, the seeds of Vigna sinensis were soaked in organic acid (100 ppm) before cultivation for 4 h and then dried for 20 h.
IAA + oxalic acid treatment: To evaluate the interactive effect of oxalic acid in combination with IAA on water stressed Vigna sinensis plants, the seeds were soaked in oxalic acid before cultivation as described before and after thinning, IAA was sprayed.
The investigated plants subjected to the different treatments were left to grow. Different growth measurements for shoot and root and photosynthetic activity of leaves as well as chemical analysis of carbohydrate, nitrogen, mineral elements as well as organic acids in shoot were carried out during the vegetative stage (45 days-old).
The data of different treatments were statistically analysed using the least significant difference (LSD) test (Snedecor and Cochran, 1980).
Analytical methods
Analysis of vegetative growth
Leaf area = Length H Maximum with H 0.75 (Turner, 1974).
Relative growth rate (RGR)
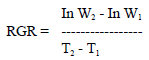 |
(Beadle, 1993), where W1 and W2 are the plant dry weights (kg) at times T1 and T2 (days), respectively.
Specific leaf area = S/W (Beadle, 1993), where S and W are the area and dry weight of plant leaves, respectively.
Net assimilation rate (NAR):
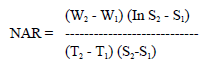 |
(Beadle, 1993), where S1 and S2 are the assimilatory areas at times T1 and T2, respectively and W1 and W2 are plant dry weights at time T1 and T2, respectively.
Photosynthetic assimilation of 14CO2: As described by Shaddad (1979), a known quantity of fresh leaf discs was introduced into a fixation apparatus designed to allow 14CO2 to be photosynthesized by the discs. The experiment was carried out under natural sun light at 10 am when light intensity was about 3500 Lux. At the end of the fixation period (10 min), the leaf discs were quickly transferred to boiling 85% (v/v) aqueous methanol for extraction of photosynthates. After cooling, the insoluble fraction was separated from the soluble one by centrifugation at 2000 g for 10 min. The soluble and insoluble fractions were made up to a volume of 35 and then 2cm3 of samples were pipetted for radioactivity measurements.
Radioactivity measurements: As described by Soliman (1984) radioactivity measurements of all liquid samples were carried out by liquid scintillation counting using a Rack Beta liquid scintillation counter (LKB-Wallac, Model 526). The counts min–1 (cmp) obtained were then calculated according to the efficiency of the apparatus used. The radioactivity measured is directly proportional to the amount of CO2 fixed in the organic compounds and this was calculated as cpm mg–1 fresh weight of leaves which was then related to dry weight.
Determination of carbohydrates: Estimation of total soluble sugars and sucrose were determined in dry powder using modifications of the procedures of Yemm and Willis (1956) and Handel (1968), respectively. While the method adopted for extraction and determination of polysaccharides in control and treated plants was that described by Naguib (1963)
Determination of nitrogenous content: Nitrogenous constituents were extracted from the dry powdered tissue by the method of Yemm and Willis (1954). Total soluble-N was determined in the extracts and total-N in the dry tissues by the conventional micro-Kjeldahl method. Subtraction of total soluble-N from total N gave the value for protein-N.
Determination of cations: For estimation of cations, dry samples were digested in a mixture of concentrated nitric acid and perchloric acid (4:1, v:v and made upto a fixed volume with deionized distilled water as described by Chapman and Pratt (1978). Flame-emission Spectrophotometry was used for determining K and Na while Ca, Mg, Mn, Fe were measured by atomic absorption spectrophotometer.
Estimation of organic acids
Oxalic acid and citric acids: The method used for estimation of oxalic and citric acid, in the present tissue extracts, was essentially that described in Snell and Snell (1949).
Keto acid: The principle of the method (Friedman and Haugen, 1943), was used for estimation of keto acids, depend on the formation of the respective hydrazones, purification of such hydrazones by extraction with suitable organic solvents and re-elution of these hydrazones in sodium carbonate solution which on alkalination would give a urine red colour that can be measured photometrically.
RESULTS AND DISCUSSION
Growth and distribution of plants are affected chiefly by too little or too much water. Expansive growth is an overriding physiological events, which can be considered as the combined results of the influences of the environmental and metabolic events on the overall plant productivity. Water stress, induced either by waterlogging or salinization affects practically every aspect of plant growth (Younis et al., 1999; Nemat Alla et al., 2001).
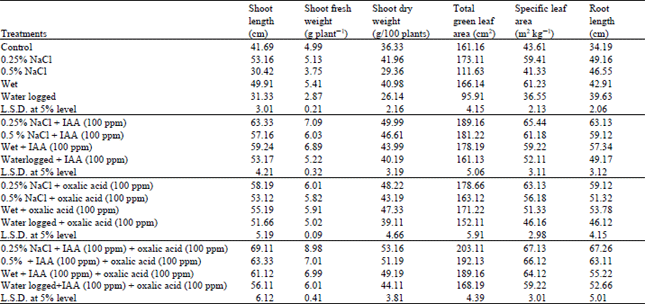
The results showed that the lowest concentration of salinity (0.25% NaCl) and the wet treatment resulted a pronounced increase in growth criteria of Vigna sinensis shoots (shoot length, fresh and dry weight, totl green leaf area and the specific leaf area) above the respective control values (Table 1). In support of this stimulation of growth criteria in response to mild water stress, Shaddad et al. (1990) recorded a promotion, rather than inhibition in dry weight production at moderate level of salinity stress. Water stress as induced by the highest level of salinity (0.5% NaCl) and the excessive supply of water (waterlogging) revealed a marked reduction in the previous growth criteria of Vigna sinensis shoot. This reduction has been documented by many authors (Kamel et al., 1995; Terbea et al., 1995; Younis et al., 1999).
Perusal of the data indicated a correlation between the reduction of dry matter under water stress conditions as induced by the highest level of salinity (0.5%) or by waterlogging treatment and the reduction which was elicited in the leaf area and the net photosynthesis (Table 1, 2).
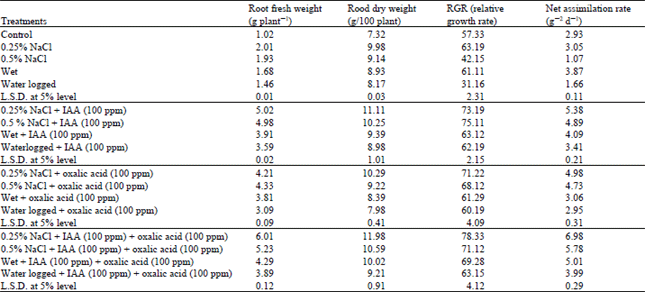
| Table 1: | Changes in growth criteria of water stressed Vigna sinensis plants treated with IAA and/or oxalic acid |
  | |
In this respect Hale and Orcutt (1987) stated that water stress substantially induces loss of turgor which affects the rate of cell expansion and ultimate cell size, consequently it decreases growth rate, stem elongation, leaf expansion and stomatal aperture.
The observed reduction in shoot growth (Table 1) which was accompanied by stimulation in growth criteria of root in response to 0.5 % NaCl and waterlogging treatments is decomended by Bradford and Hsiao (1983) and Kalapos et al. (1996), they reported that the restriction of shoot growth and continuation of root growth are both important adaptation to water stress. Since abscisic acid is a potent inhibitor of shoot growth, accumulation of abscisic acid during water stress may participate in this developmental adaptation. It is evident in this work that the relative growth and net assimilation rate were markedly reduced in Vigna sinensis plant under the highest level of NaCl (0.5%) and the waterlogging treatment, this reduction could be attributed to the reduction of net photosynthesis and green leaf area and consequently shoot dry matter production (Table 1 and 2). This conclusion is confirmed by results obtained by Younis et al. (1999) and Nemat Alla et al. (2001).
Treatment with IAA and oxalic acid either alone or incombination to water stressed plant induced an additional stimulation to the growth of wet and salinized plants at low level (0.25%) and counteracted the reductive effect of the relatively high level of salinity (0.5%) and waterlogging on shoot growth of Vigna sinensis plant comparable to the respective controls.
| Table 2: | Changes in photosynthetic ctivity (14CO2-light fixation) of water stressed Vigna sinensis leaves treated with IAA and/or oxalic acid. Values listed are expressed as cpm–3 g–1 fresh tissue min–1. |
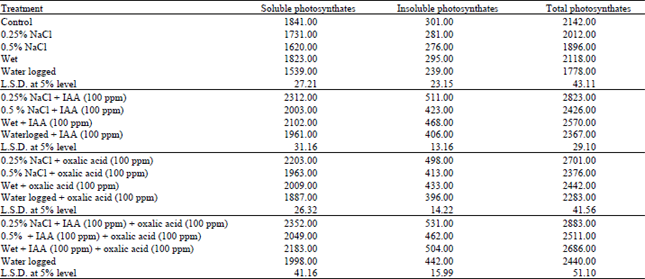 | |
| Table 3: | Changes in carbohydrate contents of water stressed Vigna sinensis shoot treated with IAA AND/OR Oxalic acid. Each value is the mean of 3 samples calculated as mg glucose equivalent g–1 dry weight. |
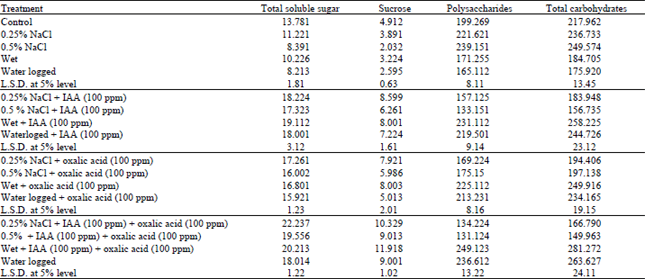 | |
Thus the values of shoot length, shoot fresh and shoot dry weight, total green leaf area and specific leaf area of different water stressed plants were lower than that of the plants treated with IAA and/or oxalic acid (Table 1). Shaddad et al. (1990), Girma and Kreig (1992) and Hamdia et al. (1996), stated that the improvement of plant growth in response to organic acids application was due to osmoregulatory role of these organic solutes which help the plants to absorb more water.
The results showed that photoassimilation of 14CO2 into soluble photosynthetic metabolites in Vigna sinensis plants were non-significantly affected under the effect of wet treatment while waterlogging treatment and salinity at the two concentrations used 0.25% and 0.5%) showed a marked reduction in this parameter below the respective controls (Table 2). The pattern of changes in insoluble photosynthetic metabolites and the total photosynthates in the tested plant appeared more or less comparable with those of soluble ones.
Application of IAAS or oxalic acid single or incombination with each other to the water stressed Vigna sinensis plant seemed to completely alleviate the deleterious effects of water stress on photosynthetic activity.
| Table 4: | Changes in nitrogen content of water stressed Vigna sinenesis shoot treated with IAA and/or oxalic acid. Values listed are expressed as mg nitrogen/100 g dry weight of shoot |
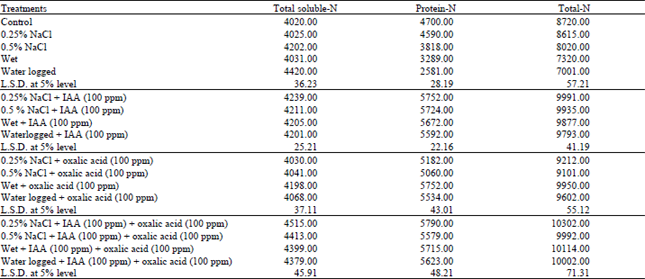 | |
| Table 5: | Changes in mineral elements of water stressed Vigna sinensis shoots treated with IAA and/or oxalic acid. Values listed are expressed as μmol g–1 dry weight |
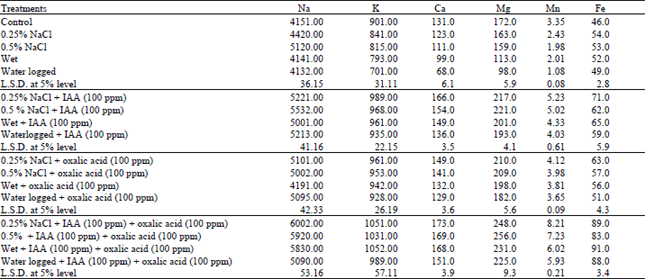 | |
These results are in agreement with those obtained by Sultana et al. (1999) using Oryza sativa, Allakhverdiev et al. (2000) using Syechoccus, they stated that excessive water supply as well as salinity significantly reduced PS1 and PS11.
Furthermore, Habib-ur-Rehman (1999) reported that waterlogging significantly reduced net photosynthesis of maize plants. These findings may suggest that excessive supply of water could inhibit photosynthesis probably by inducing stomatal closure or by affecting the photosynthetic machinery at the mesophyll and chloroplast levels. Exogenous application of IAA to the stressed plant greatly nullified the observed decline in photosynthetic activity. In this respect, Bradford (1982) found that IAA maintained the assimilative capacity of flooded plants. It became evident that exogenous application of IAA appeared to supply more or less sufficient quantities which were implicated in the recovery of growth under conditions of water stress. This recovery may be a consequence of several roles played by such a hormone which can cause triggering of the internal cellular metabolism and also induce alterations in the ratios of the growth regulators which have been shown to be critical determinators of growth and differentiation.
Mohamed and Alain (1995) and Hamdia et al. (1996) stated that organic acids counterbalanced the inhibition effect of water stress on photosynthetic activity by increasing the photosynthetic pigment and probably through more production of photosynthetic enzymes.
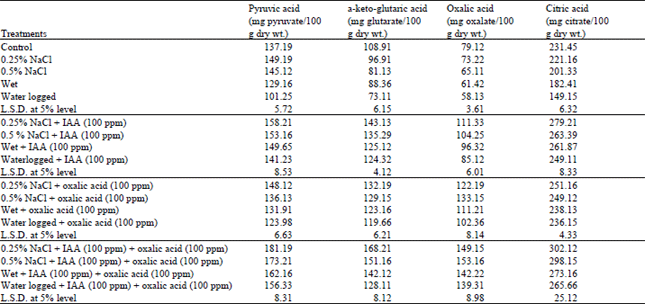
| Table 6: | Changes in Keto and organic acids of water stressed Vigna sinensis shoot treated with IAA and/or oxalic acid |
 | |
Salinity significantly decreased the soluble sugars and sucrose contents of Vigna sinensis plant as compared to the control values, this reduction was greater in case of 0.5% NaCl treatment than that observed under the effect of 0.25% NaCl treatment (Table 3). On the other hand, the reserve sugars (Polysaccharides and total carbohydrates) increased significantly under the effect of salinity at the two concentrations used (0.25% and 0.5%). In this respect Younis et al. (1993), Gao and Sagi (1998) postulated that increasing NaCl concentrations caused a progressive decrease in soluble carbohydrates in both shoot and root of Phaseolus vulgaris.
Foliar spraying of IAA at concentration (100 ppm) to the salinized plant under investigation resulted a pronounced increase in soluble sugars, whereas the insoluble sugars showed a reversible situation. In this connection Peltier and Marigo (1999) and Shukry (2001) found that the increase in soluble carbohydrates at the expense of reserve sugars was related to the conversion of insoluble carbohydrates to soluble sugars by IAA to increase the osmotic pressure. In support of this finding, El-Lawendy (1985) stated that spraying kinetin, IAA or GA3 to the plant stressed by salinity resulted in increased carbohydrate contents. In the same pattern, Abdalla et al. (1992) stated that foliar application of 10 ppm benzyl adenine, 1 ppm 2,4-D and 50 ppm GA3 increased markedly the values of reducing sugars, sucrose, polysaccharides and total sugars in leaves of red radish plants grown under stress conditions. This enhancement would eventually substantiate the amounts of soluble and insoluble sugars in the carbohydrate pool resulting in alleviation of the injurious effects induced by water stress. The decreased levels of these products of Photosynthesis could come, therefore, from a reduced efficiency of photosynthetic rate that is derived by activities of photosynthetic enzymes.
Application of oxalic acid to salinized Vigna sinensis plant appeared to compensate the reduced of soluble sugars contents caused by salinity. Shaddad et al. (1990), Girma and Kreig (1992) and El-Shahaby et al. (1994), stated that organic acids increased the organic solutes which had osmoregulatory role and consequently increased the rate of water absorption. Also Cossin and Sinha (1965) reported that organic acids are important intermediary metabolites which providing carbon skeleton for the synthesis of carbohydrate. These results are in conformity with results of Hamdia et al. (1996) and Younis et al. (1999).
These results also showed that wet and waterlogging treatments resulted a marked reduction in carbohydrate fractions (Total soluble sugars, sucrose, polysaccharides and total carbohydrates) in Vigna sinensis plants below the control levels, the rate of reduction in these fractions was most pronounced with waterlogging treatment than that observed in wet treatment (Table 3). In this connection Bradford (1982) and Younis et al. (1999) detected a decline in carbohydrate contents in response to excessive water supply. Nemat Alla et al. (2001) attributed this depression in carbohydrate pole to the inhibition in the activity of Rubisco enzyme (Ribulose-1,5-biphosphate carboxylase/oxygenase) under the effect of flooding, Bradford (1982) and De Herralde et al. (1998) stated that Rubisco enzyme inhibit the photosynthetic efficiency (reduced the activity of photosystems I and II) and decreased CO2 in the intracellular space of stomata, this reflected a reduction in carbon allocation to new leaves resulting in reduction of photon yield of CO2 assimilation and consequently limited starch synthesis in flooded plants.
Hamdia et al. (1996), Shukry (2001) and Mamdouh et al. (2002) suggested that oxalic acid and IAA increased the resistance of plants to water stress condition probably by the delay of senescence and/or on enhanced the synthesis of Rubisco.
The data revealed that water stress induced by higher level of salinity (0.5%) or waterlogging increased significantly the total soluble-N, whereas the protein and total-N were, in general, decreased under this stress conditions (Table 4). On the other hand, non-significant effect in total soluble-N in response to 0.25% NaCl and wet treatment was observed whereas a marked reduction in protein-N and total-N was elicited below the control level. In this work water stress induced by salinity or waterlogging appeared to induce more decline in protein-N which was accompanied by increments in total soluble-N is according to the inhibition of protein synthesis which occur as a result of active proteolysis or by retarded transamination processes. In this respect, Yeo (1998) concluded that the high activity of protease in water stressed plants appears to be of adaptive significance because it lead to accumulation of free amino acids as a result of the degradation of protein. The increased level of amino acid together with organic acids and quaternary ammonium compounds serve as compatible cytoplasmic solutes to maintain the osmotic balance between the cytoplasm and the vacuole under water stress conditions.
Application of IAA to Vigna sinensis plants under water stress induced by salinity (0.25% and 0.5% NaCl) and different water treatments (wet and waterlogging), increased significantly the total soluble-N, total-N as well as protein-N. The increase in total soluble-N may be due to the increase in amino acids as recorded by Shukry (2001). The protein content were increased by IAA treatment, these increments could be due to the direct increase of mRNA and protein synthesis (Singh et al., 1987). Moreover, Hathout et al. (1993) found that the protein content of tomato plants were increased significantly subsequent to foliar spray with 10 to 80 ppm IAA, particularly at the lower concentration.
Concerning the effect of oxalic acid, total soluble-N was in general, non-significantly increased under the effect of water stress induced by salinity at the two concentrations used (0.25% and 0.5%), whereas total and protein-N were stimulated under this condition, wet and waterlogging treatment showed a significant increase in these nitrogen fractions above the control levels. Combination of oxalic acid with IAA induced highly significant increase in these nitrogen fractions as compared to the respective control (Table 4). These results agree with those of many authors who recorded that organic acids are very important for plant osmotic adjustment under stress (Morgan, 1984) and regulation of pH of plant cells (Venekamp et al., 1989).
Table 5 revealed that the internal concentrations of ions in water stressed Vigna sinensis plant was markedly changed and affected greatly with water stress induced by salinity or different water treatments. The data showed that salinity treatments (0.25% and 0.5%) tend to accumulate the internal concentration of Na+ in Vigna sinensis shoot above the control levels, this increment in Na+ was more pronounced with the highest level of salinity (0.5%). On the other hand, wet and water pogging treatments non-significantly affected the Na+ in Vigna sinensis shoot. These results are in conformity with the results obtained by Abbas et al. (1991). Also the results showed that K+ concentration was in general decreased in Vigna sinensis shoot in response to water stress induced by salinity (0.25% and 0.5%) and different water treatments (wet and waterlogging). The magnitude of reduction was elicited in case of waterlogging treatment. This reduction in K+ is compatible with the results of Volaire and Thomas (1995) who reported that the reduction of K+ content in response to water stress was due to restricted transpiration or impaired ion transport. Also water stress led to significant reduction in Ca++ concentration, this depression in the uptake and transport of Ca++ under stress conditions has been reported in barley (Halperin et al., 1997).
Wolf et al. (1990) stated that the reduction in Ca++ transport under water stress conditions has been postulated to arise either from lowered transpiration rates or from inhibition of xyleme loading of Ca++. Halperin et al. (1997) suggested that the latter hypothesis and they demonstrated that symplastic transport of Ca++ is more inhibited than apoplastic transport in salinized barley plant. The concentration of Mg++ was sharply reduced in shoots of Vigna sinensis under the effect of stress condition as compared to the respective control values. Fischer et al. (1998) stated that the reduction in Mg++ is accordingly to inhibition of uptake and transport of this ion from root to shoot under stress conditions. Chemical analysis for Fe, Mn in Vigna sinensis shoot under the effect of water stress induced by salinity (0.25% and 0.5%) or different water treatments (wet and waterlogging) showed a significant reduction in these two ions below the respective control levels.
It is clear from the result that the reductive effect of water stress on metal ions content (K, Ca, Mg, Mn and Fe) of the investigated plant was alleviated by foliar application of IAA or oxalic acid either alone or incombination with each other to the water stressed plant (Table 5). The observed increase in the cations content in water stressed plants under salinity of water treatments led us to posulated that the alleviation may be related to the recovery of the plants metabolism of the elements solutes in the tissue of the shoot (Tables 2, 3, 4). This postulation is recorded by the results obtained by Ryu and Lee et al. (1996) and Shukry (2001).
The data showed that the determined keto acids (Pyruvic and α-keto-glutaric acids) were markedly decreased in shoot of Vigna sinensis plants in response to water stress induced by wet and waterlogging (Table 6). Cossins and Sinha (1965) reported that the predominant decrease in α-pyruvic and α-ketoglutaric acids which was usually accompanied by accumulation in soluble-N can be attributed to the stimulatory effect of these treatments on transamination reactions. These results are in agreement with the results obtained by Nemat Alla et al. (2001).
The data in the previous table revealed that salinity treatments (0.25% and 0.5% NaCl), in general, increase the pyruvic acid and decrease the α-ketolutaric acid content as compared to control levels. El-Shahaby et al.(1994) suggested that salinity treatments had different effects on the utilization and translocation of pyruvic and α-ketoglutaric acids within the plants.
As compared with the control, water stress induced by water logging or salinity (0.25% and 0.5% NaCI) markedly decreased the oxalic and citric acid contents in Vigna sinensis shoot. These results are in agreement with EI-Bastawisy (1999) who stated that the differences in organic acids in response to water stress conditions may be a reflection of the physical conditions within the plant cells (relative turgidity, water retention, dehydration or coagulation of protoplasm) that would lead to disturbances in metabolic activities controlled by ribosomes and enzymes. Girma and Kreig (1992) and Hamdia et al. (1996) stated that organic acids may be broken down to supply energy when anions are absorbed in excess of cations.
It is apparent from the results that application of IAA or oxalic acid to water stressed Vigna sinensis plants either alone or incombination with each other showed a remarkable increase in organic acids (oxalic and citric) and keto acids (pyruvic and α-ketoglutaric) above the control levels. This increments led us to conclude that the stimulation in synthesis of these acids inside the plant is related to the increasing in photosynthetic rate and biosynthesis of carbohydrate which utilized in synthesis of these acids to play a protective role in water stress conditions in addition to the metabolic function.
These protective functions consist in binding the excess ions absorbed by the plant and in maintaining the electrical neutrality of the cells and finally neutralizing the basic compounds (Rao and Rao, 1978). Good and Zaplachinski, (1994) reported that the increase in organic and keto-acids are important for plant osmotic adjustment under water stress and regulation of pH of plant cells.
REFERENCES
- Allakhverdiev, S.I., Y.N. Sakamota and N. Murata, 2000. Inactivation of photosystesm I and II in response to osmotic stress in Synechococcus: Contribution of water channels. Plant Physiol., 122: 1201-1208.
Direct Link - Ryu, B.Y. and J.S. Lee, 1996. Property changes in mixed media for pot flower made of several organic materials. J. Korean Soc. Hortic. Sci., 37: 127-135.
Direct Link - El-Shahaby, O.A., M.E. Younis, M.N.A. Hasaneen and A.M. Gaber, 1994. Plant growth, metabolism and adaptation in ralation to stress conditions: XIX. Effect of water stress induced by withholding and excessive water supply on carbohydrate and nitrogen metabolism in Vicia faba plants. J. Environ. Sci., 8: 91-106.
- Van Handel, E., 1968. Direct microdetermination of sucrose. Anal. Biochem., 22: 280-283.
CrossRefDirect Link - Kalapos, T., R. van den Boogaard and H. Lambers, 1996. Effect of soil drying on growth, biomass allocation and leaf gas exchange of two annual grass species. Plant Soil, 195: 137-149.
CrossRef - Mohamed, N. and C. Alian, 1995. Effect of sodium chloride on growths, tissue elasticity and solute adjustment in two Acacia nilotica subspecies. Physiol. Plant., 93: 217-224.
Direct Link - Morgan, J.M., 1984. Osmoregualtion and water stress in higher plants. Ann. Rev. Plant Physiol., 35: 299-319.
Direct Link - Nemat Alla, M.M., M.E. Younis, O.A. El-Shihaby and Z.M. El-Bastawisy, 2001. Effect of kinetin on photosynthetic activity and carbohydrate content in waterlogged or seawater treated Vigna sinensis and Zea mays. J. Biol. Sci., 1: 918-924.
CrossRefDirect Link - Peltier, J.P. and G. Marigo, 1999. Drought adaptation in Fraxinus excelsior L.: Physiological basis of the elastic adjustment. J. Plant Physiol., 154: 529-535.
CrossRefDirect Link - Timpa, J.D., J.B. Burke, J.E. Quisenberry and C.W. Wendt, 1986. Effects of water stress on the organic acid and carbohydrate compositions of cotton plants. Plant Physiol., 82: 724-728.
PubMed - Turner, N.C., 1974. Stomatal behavior and water status of maize, sorghum and tobacco under field conditions. II. At low soil water potential. Plant Physiol., 53: 360-365.
Direct Link - Venekamp, J.H., J.E. Lampe and J.T.M. Koot, 1989. Organic acids as source for drought induced proline synthesis in field bean plants (Vicia faba L.). J. Plant Physiol., 133: 654-659.
Direct Link - Chapman, H.D. and P.F. Pratt, 1978. Methods of Analysis for Soils, Plant and Water. Califorina University Division Agriculture Science Priced Publication, Califorina, pp: 50-169.
Direct Link