
Nurdilek Gulmezoglu
Department of Field Crops, Faculty of Agriculture, Osmangazi University, 26480 Eskisehir, Turkey
Pakistan Journal of Biological Sciences
Year: 2004 | Volume: 7 | Issue: 7 | Page No.: 1164-1167







ABSTRACT
The research was conducted with three triticale cultivars planted in randomized block design in three replications in 2002-2003 growing season under Central Anatolian conditions of Turkey. It was studied to estimate the grain number per spike, percentage of seed set and thousand-grain weight in spontaneous cross-pollinated, self-pollinated and untreated spike. It was determined that the seed set of spontaneous cross-pollinated, self pollinated and untreated spike varied from 57.6-64.33%, 65.36-81.80% and 84.23-88.10%, respectively. The highest thousand-grain weight amongst triticale genotypes used in the trial was found in untreated spikes (mean; 40.15 g).
PDF Abstract XML References Citation
How to cite this article
Nurdilek Gulmezoglu, 2004. Spontaneous Cross Pollination Studies on Triticale (x Triticosecale wittmack) Genotypes. Pakistan Journal of Biological Sciences, 7: 1164-1167.
DOI: 10.3923/pjbs.2004.1164.1167
URL: https://scialert.net/abstract/?doi=pjbs.2004.1164.1167
DOI: 10.3923/pjbs.2004.1164.1167
URL: https://scialert.net/abstract/?doi=pjbs.2004.1164.1167
INTRODUCTION
Triticale is a hybrid resulting from the crossing of wheat with rye and combines many of the best qualities of both of its parents. Wheat is a typical self-pollinated and is subsequently homozygous and true breedin–[1]. Though autogamy is the rule in triticale, certain tendencies toward out crossing, inherited from the rye parent, are clearly noticeable[2].
Although it is evident that there are genotypic differences between wheat varieties with regard to their ability to cross with rye, it is also certain that this ability may be influenced by environmental conditions. Yeung and Larter[3] through the use of dominant gene markers, demonstrated that cross-pollination between triticales might occur even when the plants are as much as 12 m apart. They also found that the cross-pollination rate of triticales might change dependant on the parent combination within the triticale, as well as the environmental conditions the plant was exposed to during flowering period. Rupert et al.[4] observed the growth of cultivars and lines of triticale through three generations. He found remnants of self-sterility and a tendency to out crossing in hexaploid triticale. In the first generation a significantly lower average of the seeds per spikelet rate was obtained for bagged plants (1.05) than for unbagged plants (1.73), demonstrating a high degree of out crossing in the latter. By the third generation, this difference had fallen to 1.9 and 2.0 seeds per spike obtained from bagged and unbagged plants, respectively. This indicated that the seed purity of triticale is reduced when sown repetitiously.
Due to the determinate cross-pollination nature of triticale genotypes, some research has been carried out in the field. Kiss[5] indicated that triticale was in general self-pollinating, but that cross pollination occurred at the rate of 60% in his study. Yagbbasanlar[6] and Tosun et al.[7] have also conducted research into spontaneous cross-pollination. In order to estimate the seed set, total florets per spike and total kernel per spike in the emasculated spikes of cultivars and test lines of triticale were studied, resulting in average percentages for spontaneous cross-pollination of 20.69 and 13.59%, respectively.
The aim of this study was to estimate the seed set rate in both spontaneous cross-pollinated and self-pollinated spikes of triticale cultivars. The triticale cultivars were of different physiological characters and were grown under the field conditions of Central Anatolia.
MATERIALS AND METHODS
The research was conducted on three winter triticale cultivars (Karma 2000, Presto and Tatlicak 97) during the 2002-2003 growing season. All work was carried out in EskiÕehir, a province in the Central Anatolian region of Turkey.
Sowing was carried out on the 15th of October, 2002. The experiment was designed to measure the differences in the seed sets of cross-pollinated, self-pollinated and intact spikes. Plots (6 m2) consisted of four 3 m rows seeded 30.5 cm apart, with a sowing density of 350 viable grains per square meter in a random block design. The plots were replicated three times. Four rows of the triticale cultivars used in the experiment were sown in the field at a distance of two meters from the trait area in order to determine the seed set by cross-pollination, under field conditions. Phosphorus (P2O5) and Nitrogen (N) were applied at the time of p lanting, at a standard rate of 60 kg ha–1 P2O5 and 23.40 kg ha–1 N (the fertilizer applied was di-ammonium phosphate: 18-46-0). Nitrogen topdressing was then applied during the tillering stage in March; this being composed of ammonium sulphate (21-0-0) completed to total 70 kg N ha–1. Annual precipitation, average temperature and humidity rates (from October 2002 to July 2003) were 297.1 mm, 8.6°C and 62.3%, respectively. The average rates of precipitation, temperature and humidity recorded during the month of May, when treatment was carried out, were 43.5 mm, 16.4°C and 53.2%, respectively. Soil samples (0-30 cm) were taken before planting and analysed for some parameters. The soil of the experiments was determined as loam, low organic matter, moderate level of CaCO3 and no salinity problems.
The data were tested for analysis of variance using the statistical package, MSTAT-C[8] and the differences between means were compared by the Least Significant Differences. Statistical significance was declared at P<0.01.
The emasculated flowers: Fifteen spikes were selected from each triticale cultivar, about 3 to 4 days before expected flowering (Karma 2000 on May 14th, Presto and Tatl2cak 97 on May 21st 2003). Four or five spikelets were cut off from both the bottom and top of the spike and 20 spikelets from the centre of each spike were retained. The centre florets of each spikelet were pulled out due to the immaturity of the ovaries. Scissors were used to cut the glumes, lemma and palea from just above the top of the stigma and in this way no damage occurred to the stigma or anthers. The three anthers were then carefully removed from each floret with forceps for the purpose of cross-pollination.
The self-fertile spikes: Fifteen spikes were selected at random before full exertion from the flag leaf. The selected spikes did not undergo any treatment. A bag was placed over the spike, which was then folded bottom edge over and a paperclip was placed over the fold to prevent the bag from being blown away in wind. The bagged spikes were maintained in a condition conducive for self-pollination until the milk stage. The bagged self-pollinated spikes were collected during harvesting.
Intact spikes: Fifteen random spikes in each of the plots were selected in order to determine the seed set rates for treated spikes and intact spikes after harvest.
The number of seed per spike: The first two florets of ten centre spikelets (a total of 20 florets) on each spike were counted for the three treatments.
Percentage of seed set: The seed set was calculated using the below mentioned formula;
![]()
Thousand-grain weight: Obtained seeds from treated and untreated triticale spikes were weighed after counted and thousand-grain weight was calculated.
RESULTS
The number of seeds per spike: All the genotypes of triticale reacted very differently, with significant differences in the genotypes (P<0.01), treatment (P<0.01) and genotypes x treatment interaction (P<0.01) (Table 1). The cross-pollination of the seed set per spike ranged from 11.52 to 12.87. While Presto yielded the highest seed set (12.87), Tatl2cak 97 and Karma 2000 showed similar results (11.52 and 11.7, respectively). The seed set per spike (mean 14.61) under conditions of self-pollination was higher than that in cross-pollination. In terms of self-pollination, the highest count was found in Karma 2000 and the lowest in Presto. The seed set of untreated spikes, cross-pollinated under normal conditions, was higher than that of the other treated spikes (mean 17.32). Untreated Karma 2000 demonstrated a higher seed set (17.5), while Presto produced the lowest (16.85).
Percentage of seed set (%): Owing to the percentage of the seed sets having been determined from spikes that had undergone treatment, data is shown in parallel due to its appearance in this form (Table 1). The differences between the genotypes (P<0.01), treatments (P<0.01) and genotypes x treatments (P<0.01) were significant. The percentages for seed set on cross-pollinated spikes ranged from 57.62% (Tatl2cak 97) to 64.33% (Presto). The mean cross-pollination rate between triticale genotypes was 60.15, while self-pollination was found to be highest in Karma 2000 (81.80%), followed by Tatlicak 97 (72.03%) and Presto (65.36%). According to the findings of the research, triticale genotypes crossed mostly via self-pollination. The highest rates for seed set in all the triticale genotypes were found to be those of the intact spikes; 88.10, 87.5 and 84.23 for Tatlicak 97, Karma 2000 and Presto, respectively.
| Table 1: | The seed set number per spike and seed set percentage of triticale genotypes |
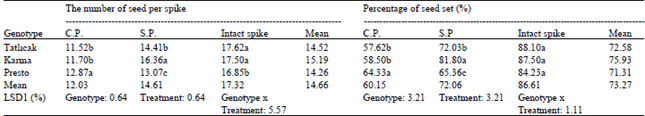 | |
Means in a column followed by the same letter(s) are not significantly different at 1 % probability, C.P., cross-pollinationS.P., self-pollination; | |
| Table 2: | The Thousand-grain weight of triticale genotypes |
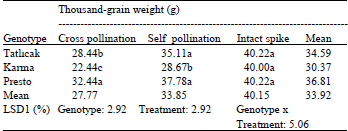 | |
Means in a row followed by the same letter(s) are not significantly different at 1% probability | |
The thousand-grain weight: The differences between the genotypes (P<0.01), treatments (P<0.01) and genotypes x treatments (P<0.01) were significant (Table 2). Of the triticale genotypes investigated, the intact spikes yielded the highest thousand-grain weight. In this research, results amongst the triticale genotypes were found to be similar (mean 40.15 g) to each other. The lowest thousand-grain weight was obtained from the cross-pollinated samples. The thousand-grain weight obtained from the self-pollination samples of triticale changed in relation to the genotypes. While Presto produced the highest thousand-grain weight, Karma 2000 produced the lowest.
DISCUSSION
In this study, significant statistical differences were obtained all for the characteristics (the number of seed set per spike, percentage of seed set and thousand-grain weight) in terms of the genotypes, treatment and genotype x treatment interaction.
The findings of the research were that seed sets were low in the cross-pollinated spikes (60.15%), while those of the intact spikes were found to be the highest. In crossed triticale, a plant is produced with the self-pollination properties of the parent wheat plant; while at the same time it possesses the cross-pollination properties of the rye parent. Although triticale is generally a self-pollinating plant, some researchers have reported cross-pollinating genotypes[3-5,9]. Kiss[5] accounted that the cross-pollination of triticale occurred at a range of between 0-60%, while Yeung and Larter[3] saw a rate of approximately 5%; Malik[10], 17.58%; Yagbasanlar[6], 20.69% and Tosun et al.[7], 13.9%. The results of this study were found to be similar to those of Kiss[5]. It is believed that the higher rates of cross-pollination recorded in these findings compared to those of some of the others may be accountable to the different genotypes and seasons in which the research were carried out. The emasculation ensures the cross-pollination of flowers on the spikes and this is the reason for the increase in the seed sets.
The rates for self-pollination in the triticale genotypes were found to be quite high. In particular, the results found in the seed sets for self-pollinating Karma 2000 were close to the results for its spikes not receiving treatment. It is believed that the one-week earlier flowering of Karma (May 14th 2003) as opposed to that of the other varieties (May 21st) may have been influenced in this way. The lowest self-pollinated seed set rates were reported in the Presto variety, although cross-pollinated seed set quantities were higher.
The decreases in the thousand-grain weights amongst the treatment spikes may have been due to their being grown under stress conditions, while no significant differences were observable amongst the thousand-grain weight falls of the intact spikes. In fact, the seed counts of the self-pollinating genotypes increased in opposition to the rate of the thousand-grain weights. The most manifest was seen in Karma 2000 with high seed counts obtained in self-pollination and thousand-grain weight falls.
In conclusion, the cross-pollination, self-pollination and intact rates of triticale genotypes used in the trials varied from 57.6-64.33, 65.36-81.80 and 84.23-88.10%, respectively. It has been seen that in order to maintain the genetic purity of the genotypes from cross-pollination, an isolation distance should be preserved around the varieties. It is suggested that the early flowering, high self-pollination rates and low cross-pollination rates of Karma 2000 seen in the trials conducted, indicate it could be preferable genotype with high thousand-grain weight and seed set rate, followed by Tatlicak 97.
ACKNOWLEDGMENTS
I would like to thank Prof. Dr. E. K2nac2 and Assoc. Prof. Dr. G. K2nac2 for scientific advice and Res. Assist. Z. Budak for assistance in this investigation.
REFERENCES
- Muntzing, A., 1979. Triticale Results and Problems. Verlag Paul Parey-Berlin and Hamburg, Germany.
Direct Link - Kiss, A., 1970. Spontaneous crossing between hexaploid triticale rosner and triticale 64. Wheat Infrom. Service, 31: 24-25.
Direct Link