
Rebeca Isabel Vergara-Reyes
Laboratory of Cell Biology, School of Veterinary Medicine and Animal Science, University of Veracruz, Veracruz, Mexico
Patricia Cervantes-Acosta
Laboratory of Molecular Biology, School of Veterinary Medicine and Animal Science, University of Veracruz, Veracruz, Mexico
Antonio Hernández-Beltrán
Laboratory of Functional Alterations, School of Veterinary Medicine and Animal Science, University of Veracruz, Veracruz, Mexico
Manuel Barrientos-Morales
Laboratory of Reproductive Biology, School of Veterinary Medicine and Animal Science, University of Veracruz, Veracruz, Mexico
Belisario Domínguez-Mancera
Laboratory of Cell Biology, School of Veterinary Medicine and Animal Science, University of Veracruz, Veracruz, Mexico
LiveDNA: 52.34940
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 3 | Page No.: 297-309







ABSTRACT
Background and Objective: Arcuate nucleus (ARC), a component of appetite-regulatory factors, contains populations of both orexigenic and anorexigenic neurons and one of the fundamental components of its system is leptin. Studies have evidenced the critical neurotrophic role in the development of ARC. To determine such effects on neuron development, N1E-115 neuroblastoma cells were used as an ARC model. Materials and Methods: N1E-115 neuroblastoma cells were treated with leptin [10 nM] for 24, 48 and 72 hrs. Dimethyl sulfoxide (DMSO) 1.5% was used as a known drug that promotes neurite expression. Cells percentage (%) that developed neurites was evaluated by bright field microscopy. Patch-clamp electrophysiology was used to analyze membrane ion currents, RT-PCR for quantifying changes in mRNA expression of anorexic peptides, proopiomelanocortin (POMC) and cocaine and amphetamine-related transcript (CART), in addition to principal Nav, Cav ion channel subunits. Results: N1E-115 cells treated with leptin show neurite expression after 24 hrs of treatment, similar effects were obtained with DMSO. Leptin (time-dependent) increases the inward current in comparison with the control value at 72 hrs. Outward currents were not affected by leptin. Leptin and DMSO increased Na+ and Ca2+ current without changes in the kinetic properties. Lastly, leptin promotes an increase in mRNA level expression of transcripts to POMC, CART, Nav1.2 and Cav1.3. Conclusion: Leptin chronic treatment promotes neurite expression, Up-regulation of Na+ and Ca2+ ion channels determining neuronal excitability, besides increasing the mRNA level expression of anorexic peptides POMC and CART in neuroblastoma N1E-115.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Rebeca Isabel Vergara-Reyes, Patricia Cervantes-Acosta, Antonio Hernández-Beltrán, Manuel Barrientos-Morales and Belisario Domínguez-Mancera, 2021. Leptin Chronic Effect on Differentiation, Ion Currents and Protein Expression in N1E-115 Neuroblastoma Cells. Pakistan Journal of Biological Sciences, 24: 297-309.
DOI: 10.3923/pjbs.2021.297.309
URL: https://scialert.net/abstract/?doi=pjbs.2021.297.309
DOI: 10.3923/pjbs.2021.297.309
URL: https://scialert.net/abstract/?doi=pjbs.2021.297.309
INTRODUCTION
The functioning of the mammalian nervous system is regulated by networks and interconnections of nerve cells and its’ dynamics depend on both the electrophysiological intrinsic properties of these networks and synaptic currents1. One of these circuits is the neuroendocrine system, which regulates food intake and energy balance2. A fundamental component of this system is the hormone leptin, which is released by white adipose tissue3. This 16 kDa cytokine plays a prominent role in the regulation of energetic metabolism and appetite4,5. Although there are complex central mechanisms of energy homeostasis, appetite is primarily controlled by circuits of hypothalamic and hindbrain nuclei, which receive inputs from central (cerebrum) and peripheral sources, such as stomach (Ghrelin), adipocytes (Leptin), pancreas (Insulin), liver (growth factors) and others6,7. The arcuate (ARC) nucleus, is a hypothalamic center associated with the regulation of energy balance and is a critical locus for food intake regulation as it integrates signals from the brainstem and the periphery8. The first-order neurons of the ARC are in direct contact with peripheral satiety factors which connect to the second-order neuron centers of dorsomedial, paraventricular and ventromedial nuclei9. Two populations of neurons controlling appetite in the ARC nuclei: One of them neurons coexpressing Neuropeptide Y (NPY) and agouti-related protein (AgRP) and the other neurons coexpressing pro-opiomelanocortin (POMC) and cocaine and amphetamine-regulated transcript (CART). NPY and AgRP stimulate food intake “Orexigenic effects”10, while POMC and CART repress it “Anorexigenic effects”11. Both orexigenic and anorexigenic functions must be developed in utero to prepare the newborn for life, studies have demonstrated a critical neurotrophic role of leptin in the development of ARC axons pathways12,13. An altered hormone or neurotrophic environment may alter critical neurodevelopmental signaling13,14. Many hormones, neurotransmitters and their receptors have been implicated in the regulation of food intake, these inputs information the brain of the metabolic status of the organism by acting on various hypothalamic (ARC) and extra hypothalamic structures. One model that has received attention involves two groups of neurons in the ARC nucleus. One subset of neurons double-stains POMC and CART, whose gene products are anorexigenic effects11, while another group co-expresses AGRP and NPY, whose gene products are orexigenic effects10, the neural activity of these groups of cells is likely influenced by metabolic information carried out by peripheral signals, such as leptin. The ARC nucleus is a region that is rich in leptin receptors15. Besides, studies have shown that leptin can regulate levels of CART/POMC or AGRP/NPY gene expression11,16. The comprehension of cellular function has been benefited by the use of numerous cell lines used such as in vitro models, in the case of the study of neuronal function and development, one cell line that has been used is the N1E-115 neuroblastoma cell line17, which is composed of undifferentiated cells, capable of unlimited proliferation when it is cultured in vitro without any stimulation. These cells are capable of developing electrical excitability and expressing neurites upon exposure to different physical or chemical agents, such as Dimethyl sulfoxide (DMSO)18,19. Many protein signaling is involved in the mechanisms leading to neuronal differentiation in N1E-115 cells20. N1E-115 cells contain mRNA for POMC, CART, AGRP, long-isoform leptin receptor (Ob-Rb) and insulin receptor (INS-R), it confers a definite “ARC-like” quality to N1E-115 cells21.
The study aims to determine the trophic (ion currents) and differentiation (neurite presence) effects of chronic treatment with leptin on “ARC-like” N1E-115 cells and to examine the level mRNA expression of specific anorexigenic neuropeptides POMC and CART.
MATERIALS AND METHODS
Study area: The research project was carried out in the Laboratory of Cell Biology, School of Veterinary Medicine and Animal Science, University of Veracruz, Veracruz, Mexico. This research project was conducted from September, 2018-July, 2020.
Cell culture: Mouse (Mus musculus) neuroblastoma cell line N1E-115 (ATCC® CRL2263)17 were kept in culture, in an RPMI 1640 medium (BioWest, NW Business Park, MO, USA), supplemented with 10% fetal bovine serum (BioWest, NW Business Park, MO, USA), L-glutamine and 1% penicillin-streptomycin (100 U mL–1) (G3126, P0781, Sigma Aldrich, St. Louis Miss, USA). Cells were incubated (Fisher Scientific, Waltham, Mass, USA) in a humid atmosphere of 5% CO2-95% air at 37°C. The incubation medium was changed every 2 days. Cells were harvested once per week by treatment with a trypsin-EDTA solution and reseeding was carried out at 20% of the original density in culture flasks (25 cm2) or in 35 mm Æ diameter Petri dishes for treatment (Table 1) with leptin [10 nM] (American Peptide Co, Sunnyvale, USA ) and/or DMSO (D2650, Sigma Aldrich, St. Louis Miss, USA).
Electrophysiology: The neuroblastoma cell line N1E-115 was subjected to electrophysiological recording using the patch-clamp technique, in the whole-cell configuration.
| Table 1: Culture medium components | |||||
| Experimental conditions | RPMI (%) | SFB (%) | Antibiotic (%) | Leptin (nM) | DMSO (%) |
| Control | 88 | 10 | 2 | 0 | 0 |
| Leptin | 88 | 10 | 2 | 10 | 0 |
| DMSO | 96 | 0.5 | 2 | 0 | 1.5 |
| Leptina+DMSO | 96 | 0.5 | 2 | 10 | 1.5 |
100% = 100 mL, Roswell park memorial institute (RPMI) 1640 medium (BioWest®), FBS: Fetal bovine serum (BioWest®), DMSO: Dimethyl sulfoxide (Sigma Aldrich®) | |||||
| Table 2: Recording solutions | ||||||||||||||
| Solution | NaCl | KCl | CaCl2 | TEA-Cl | BaCl2 | CsCl | MgCl2 | ATP/GTP | CdCl2 | TTX | EGTA | Hepes | Glu | pH |
| Totale | 145 | 5 | 5 | 0.5 | 10 | 5 | 7.3 (NaOH) | |||||||
| Nae | 150 | 2 | 0.5 | 10 | 5 | 7.3 (NaOH) | ||||||||
| Bae | 133 | 10 | 10 | 0.001 | 10 | 5 | 7.3 (NaOH) | |||||||
| Totali | 30 | 100 | 1 | 2 | 2/0.05 | 10 | 10 | 5 | 7.3 (KOH) | |||||
| Nai | 30 | 1 | 100 | 2 | 2/0.05 | 10 | 10 | 5 | 7.3 (CsOH) | |||||
| Bai | 30 | 1 | 100 | 2 | 2/0.05 | 10 | 10 | 5 | 7.3 (CsOH) | |||||
Totale, Nae, Bae, bath solutions. Totali, Nai, Bai, pipette solutions. Units are in mM, TTX: Tetrodotoxin, ATP/GTP: Adenosine triphosphate/Guanosine-5'-triphosphate, EGTA: Ethylene glycol tetraacetic acid, Glu: Glutamic acid, TEA-Cl: Tetraethylammonium chloride, Hepes: N-2-hydroxyethylpiperazine-N-ethanesulfonic acid, BaCl2: Barium chloride, CsCl: Caesium chloride, NaCl: Sodium chloride, KCl: Potassium chloride, CaCl2: Calcium chloride, MgCl2: Magnesium chloride | ||||||||||||||
Voltage-clamp was used to dissect total current components as previously described22. Recordings were carried out at room temperature (~22°C) and micropipettes were made of borosilicate glass tubing (P-30 Vertical Micropipette Puller, Sutter Instrument, Novato, CA, USA). Recording micropipettes had resistances of ~3.5-4.5 MΩ once filled with the internal recording solution. Voltage-clamp recordings were performed using an Axopatch 200B amplifier (Molecular Devices LLC. San Jose, CA, USA). After reaching the whole-cell configuration, currents were evoked by depolarizing pulses of variable amplitude and duration from a holding potential (Vh) of -80 mV applied at intervals of 1 sec, using a Digidata 1440A interface (Molecular Devices LLC. San Jose, CA, USA), controlled by a computer. Current signals were acquired at 10-100 μs (Acquisition frequency for each protocol were: Total currents, sampling rate 10.00 kHz and interval of 100.00 μs, Sodium current, sampling rate 66.67 kHz and interval of 15.00 μs and Calcium current sampling rate 10.00 kHz and interval of 100.00 μs) and signals were filtered at 5 kHz. Data was stored on the computer and analyzed using pCLAMP v10.1 (Molecular Devices LLC. San Jose, CA, USA) and Sigma Plot v11.0 (Systat, San Jose, CA, USA) software. Leakage and capacitive currents were digitally subtracted off-line. Series resistance was not compensated but cells were excluded from further analysis if values changed by >20% in the course of the recording. Membrane capacitance (Cm) and Membrane resistance (Rm) were determined as described previously23. To block INa, 1 μM TTX was added to the external solution, 0.5 mM CdCl2 was added to block ICa and 100 mM CsCl was added to block IK to the internal solution. The compositions of the recording solutions are given in Table 2. Osmolarity was adjusted to 300 and 290 mosmol L–1 for a bath (external) and pipette (internal) recording solutions, respectively. Control and treated N1E-115 cells were kept with a peptide-free culture medium for ~60 min before electrophysiological recording, to avoid the acute effects of the drugs.
Electrophysiology data analysis: Data were analyzed and plotted through the combined use of the pCLAMP v10.1 and Sigma Plot v11.0 software. Two-way analysis of variance (ANOVA) was used to assess the effect of leptin and DMSO on N1E-115 neuroblastoma cells differentiation and ion currents therein, followed by Tukey multiple comparison tests for possible pairwise differences. A p-value of <0.05 was considered to be statistically significant. Data are presented as means±S.E. unless otherwise specified. The presence of leptin was considered first and the presence of DMSO the second factor of comparison. Curve fits of the ion currents kinetics were made using the nonlinear, least-squares fitting procedure included in the Sigma Plot v11.0 software. The number of analyzed cells is given in the error bar plot. Conductance (GNa) was calculated to analyze the activation kinetics of INa+, from I-V curve values and normalized to its maximal value. The conductance of treated cells was normalized to their corresponding control. Data points in the G-V curves were fitted with the Boltzmann equation24 of the form:
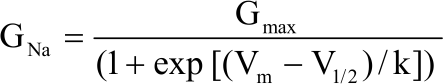 | (1) |
Where, GNa is the sodium conductance, Gmax is the maximum sodium conductance, Vm is the membrane potential, V1/2 is the voltage where 50% of the sodium channels are open and k is the slope factor which determines the shape of the activation curve.
| Table 3: Primer sequence used in the PCR reactions | ||||
| Locus | Sequence (5'-3') | bp. | GeneBank | References |
| POMC | FW 5’-GTGTTTCCTGGCAACGGAGATG-3’ | 326 | AH002232.2 | Roth et al.21 |
| RV 5’-CGTTCTTGAAGAGCGTCACC-3’ | ||||
| CART | FW 5’-CATCTACTCTGCCGTGGATG-3’ | 384 | NC_005101.4 | Roth et al.21 |
| RV 5’-GGAAAGAATTGCAGGAAGTTCC-3’ | ||||
| Nav 1.2 | FW 5’-GACGACGAAAATGGCCCAAA-3’ | 211 | NM_018733 | Baroni and Moran28 |
| RV 5’-GGTTTACAAATTCTGTTACATACGC-3’ | ||||
| Cav 3.1 | FW 5’-TTCTTCAAGGACAGGTGGAA-3’ | 355 | XR_385353 | Calderón-Rivera et al.29 |
| RV 5’-GCCTCCTTGTTGCTCTCCTC-3’ | ||||
| GAPDH | FW 5’-TCAACGGCACAGTCAAGG-3’ | 470 | XM_017321385.2 | Church et al.30 |
| RV 5’-GAGTACTTGCGCTCAGGAGG-3’ | ||||
| POMC: Pro-opiomelanocortin, GAPDH: Glyceraldehyde 3-phosphate dehydrogenase, CART: Chimeric antigen receptor T | ||||
To analyze the voltage dependence of steady-state inactivation of INa+, currents were recorded after 1s. Conditioning pulses, applied from a Vh of -100 mV, in 10 mV steps, followed by a 10-ms test pulse to -10 mV. The normalized data were plotted against the conditioning potentials. Data points in inactivation curves were fitted with the Boltzmann equation24 of the form:
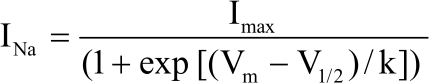 | (2) |
Where, INa is the sodium current, Imax is the maxim sodium current at test pulse, Vm is the membrane potential, V1/2 is the voltage where 50% of the sodium channels is inactivated and k is the slope factor which determines the activation curve shape.
To analyze the activation kinetics of ICa2+, tail current decay was fitted according to the method of Matteson and Armstrong25. In brief, the first step consisted of fitting a least-squares exponential to the slow component of the tail current and extrapolating this exponential to the beginning of the step. The fitted slowly exponential was then subtracted out and the remaining current was fitted with a least-squares exponential. To minimize possible errors, the initial amplitude of the tail current was taken as the amplitude of the fitted exponential, measured 100 μs after the onset of repolarization26,27. The current at each test potential was estimated by measuring the tail current amplitude. I-Voltage (I-V) curve for activation was fitted with a Boltzmann equation24 of the form:
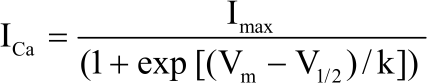 | (3) |
Where, ICa is the calcium current, Imax is the maximum tail current, Vm is the membrane test potential, V1/2 is the potential for half-maximal activation of Imax and k is a slope factor.
CART, POMC, Nav, Cav mRNA expression: Neuroblastoma N1E-115 cells were treated with leptin [10 nM] for 72 hrs, simultaneously with their respective control group. The mRNA was isolated with the Quick-RNATM MiniPrep Kit (Catalog R1054. Zymo Research, Orange, CA, USA). In summary, the method consisted of recovering and lysing the cells with the ZR RNA Buffer, subsequently, the lysate was transferred into a column in a collection tube and centrifuged at 12,000 g for 1 min. The Total RNA was purified with RNA Pre-Wash Buffer, RNA Wash Buffer and DNase/RNase free water. Eluted RNA was stored at <20°C until its analysis. RNA quality was measured through spectroscopy (Bio Photometer Eppendorf Model 6131, Hamburg, Germany), samples were accepted when integrity A260 nm/A280 nm >1.6 and RNA concentration >3 μg/10 μL. A copulated retro-transcription was applied to the mRNA with the commercial kit High Capacity cDNA Reverse Transcription (Applied Biosystems, Foster City, CA, USA), following kit protocol. Primers (Table 3) were used for measuring the gene expression as directed by the Pyrostart fast PCR Master Mix kit (Fermentas, Thermo Fisher Scientific, Waltham, Mass, USA). The PCR conditions included an initial denaturation step at 94°C for 5 min, followed by a cyclic reaction, consisting of denaturation at 94°C for the 30 sec, annealing at 55°C for 30 sec and extension for 1 min at 72°C. PCR products were then subjected to 1% agarose gel electrophoresis and nucleic acid bands were visualized through ethidium bromide staining. GAPDH was used as an endogenous control for gene expression, The Image-J® software was used for semi-quantitative analysis. Data are expressed as means±SE. Statistical significance was established using Student’s t-test (p<0.05) when two means were compared.
RESULTS
Morphological changes promoted by leptin in the N1E-115 neuroblastoma cell line: Firstly, N1E-115 neuroblastoma cells were reviewed every 24 hrs from the beginning of treatment (day 0), up to 72 hrs (day 3), through bright field light microscopy (Leica DM IL LED, Wetzlar, Germany), to analyze cell differentiation over time in each experimental conditions.
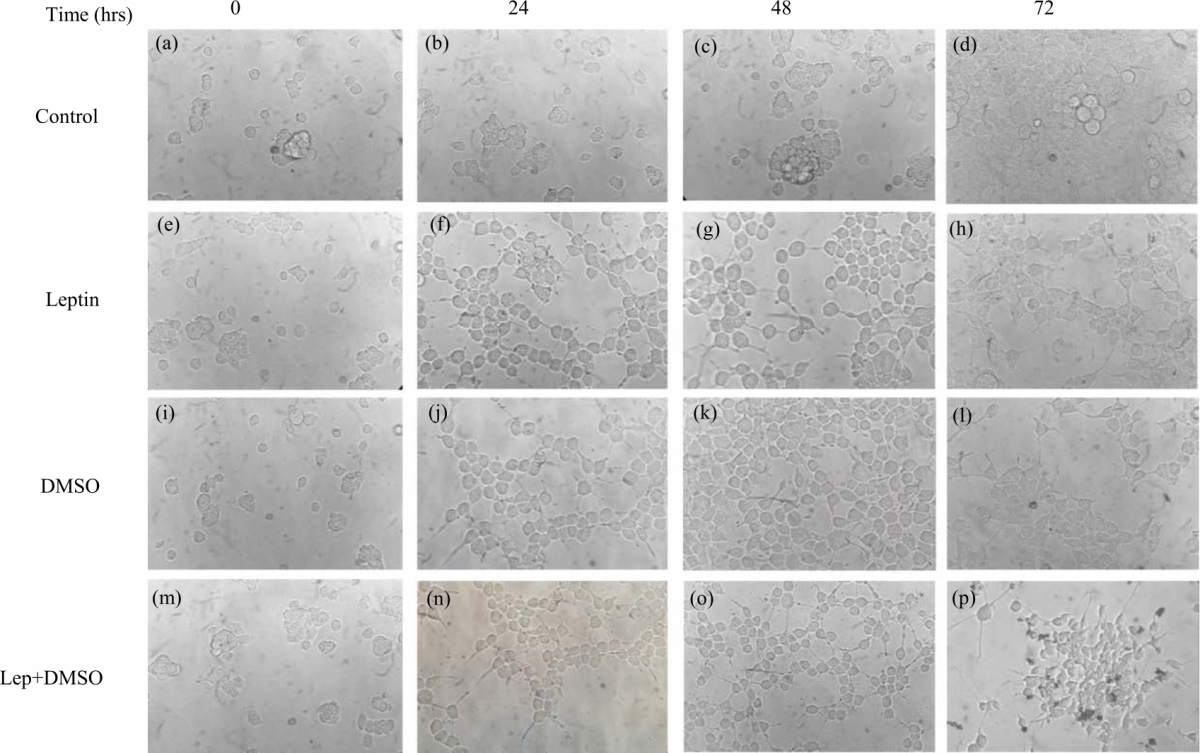 |
| Fig. 1(a-p): | Differentiation of neuroblastoma N1E-115 cell line treated with leptin or DMSO (a-d) N1E-115 cells under control condition. N1E-115 cells were exposed to different drugs (0, 24, 48 and 72 hrs), (e-h) Leptin [10 nM], (i-l) DMSO [1.5%] with FBS [0.5%], (m-p) Leptin [10 nM]+DMSO [1.5%] with FBS [0.5%] |
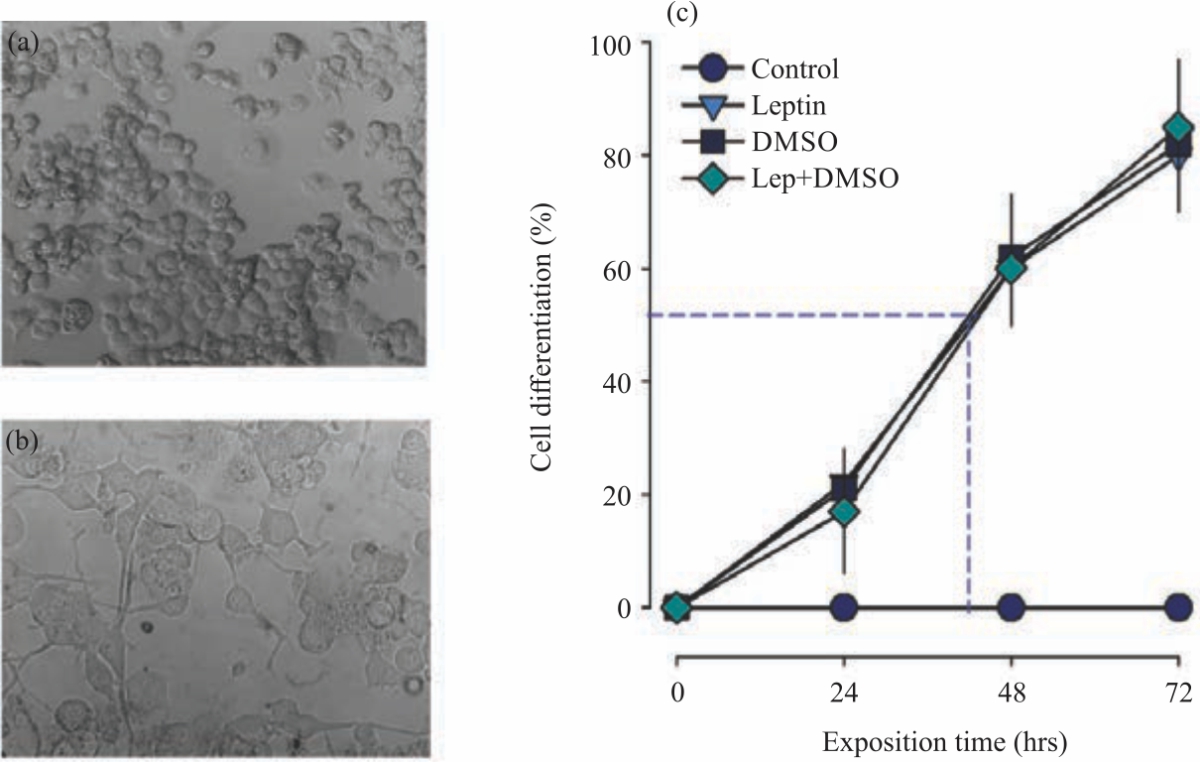 |
| Fig. 2(a-b): | Leptin promotes differentiation of neuroblastoma N1E-115 cell line (a) Neuroblastoma N1E-115 cells under control conditions (20X), (b) Differentiated N1E-115 cells treated with leptin [10 nM] for 72 hrs (40X), (c) Time course of cells differentiated in different treatments, the number of differentiated cells per optical field (n = 10 optical fields per day per treatment) was quantified and divided by the number of cells. The blue short dashed line indicates 50% differentiated cells and the time over which it occurs |
It should be mentioned that DMSO was used to add a positive control to the experiment and to show the cell differentiation promoted by a known drug. In Fig. 1a, d, it can be observed that the N1E-115 cells do not modify their morphology over time (0, 24, 48 and 72 hrs) under control conditions. Changes in the morphology can be appreciated in N1E-115 neuroblastoma cells when they are chronically (0, 24, 48 and 72 hrs) treated with leptin (Fig. 1e-h), the appearance of neurites (axon and dendrites) is observed after 24 hrs of treatment, in a similar way as can be seen in the treatment with DMSO (Fig. 1i-l). Treatment with Leptin plus DMSO (Fig. 1m-p) similarly promotes cell differentiation as can be seen with each drug, separately.
The proportion of cell differentiation per day was established to calculate the number of differentiated cells per optical field. The number of differentiated cells was quantified through a bright field light microscope and divided by the total of cells, thus showing a proportion of cells that change their morphology through drug exposition time, this quantification is observed in Fig. 2a, b. In a time close to 48 hrs (Fig. 2c), 50% of cells show differentiation, within 72 hrs, more than 80% of cells have developed morphological changes (neurites). Leptin, DMSO and leptin plus DMSO similarly promote cell differentiation (p>0.05).
Biophysical changes promoted by leptin in N1E-115 neuroblastoma cell line: One aspect that was decided to evaluate was whether the morphological changes were accompanied by changes in cell biophysical properties. Neuroblastoma N1E-115 cells show inward and outward ion currents under control conditions, that is, without the use of drugs to promote their differentiation (Fig. 3a, b). Neuroblastoma N1E-115 cells, when exposed to leptin, DMSO and leptin plus DMSO (72 hrs), increase their inward ion currents (Fig. 3a-c), this increase showed in chronically treated cells with Leptin and DMSO was significant (p<0.05), moreover depends on exposition time (Fig. 3d). Increments in outward currents occurred only in cells treated with DMSO (Fig. 3e, f) and depend on exposition time (Fig. 3g).
Leptin effects on INa+ in neuroblastoma cell line N1E-115: It was decided to carry out voltage-clamp experiments to analyze whether chronic treatment with leptin, DMSO or Leptin plus DMSO, increased any of the inward current components, sodium or calcium current or both. Neuroblastoma N1E-115 cells show fast activation (1-2 ms) and fast inactivation (2-4 ms) sodium currents with an amplitude ~2 nA (2000 pA) (Fig. 4a) in control conditions.
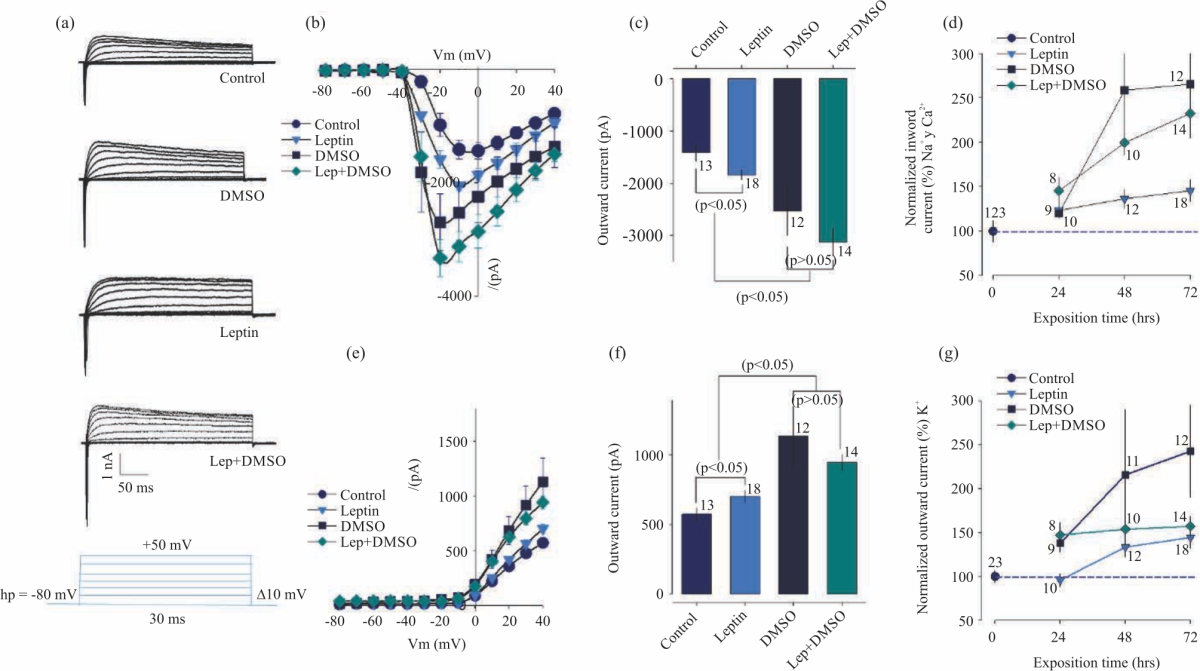 |
| Fig. 3(a-g): | Chronic treatment with leptin or DMSO enhances membrane currents in neuroblastoma N1E-115 cells (a) Representative whole-cell patch-clamp family currents recorded in response to 300 ms command steps ranging from -80-+50 mV, from a holding potential (hP) of -80 mV in control and treated cells, (B) Current-voltage relations of inward currents recorded from cells as in A, peak inward current was measured within 1-2 ms of the onset of voltage command, (c) Bar chart summarizing the effects of Leptin or DMSO on inward currents recorded from cells as in A, (d) Time course dependent of stimulatory action of leptin or DMSO on inward currents; currents were converted to a percentage of its average value in control cells, (e) Current-voltage relations of outward currents recorded from cells as in A, outward current was measured at the end of the voltage command (295 ms), (f) Bar chart summarizing the effects of Leptin or DMSO on outward currents recorded from cells as in A and (g) Time course dependent of stimulatory action of leptin or DMSO on outward currents, currents were converted to a percentage of its average value in control cells. *Significant differences (p<0.05) compared with control cells |
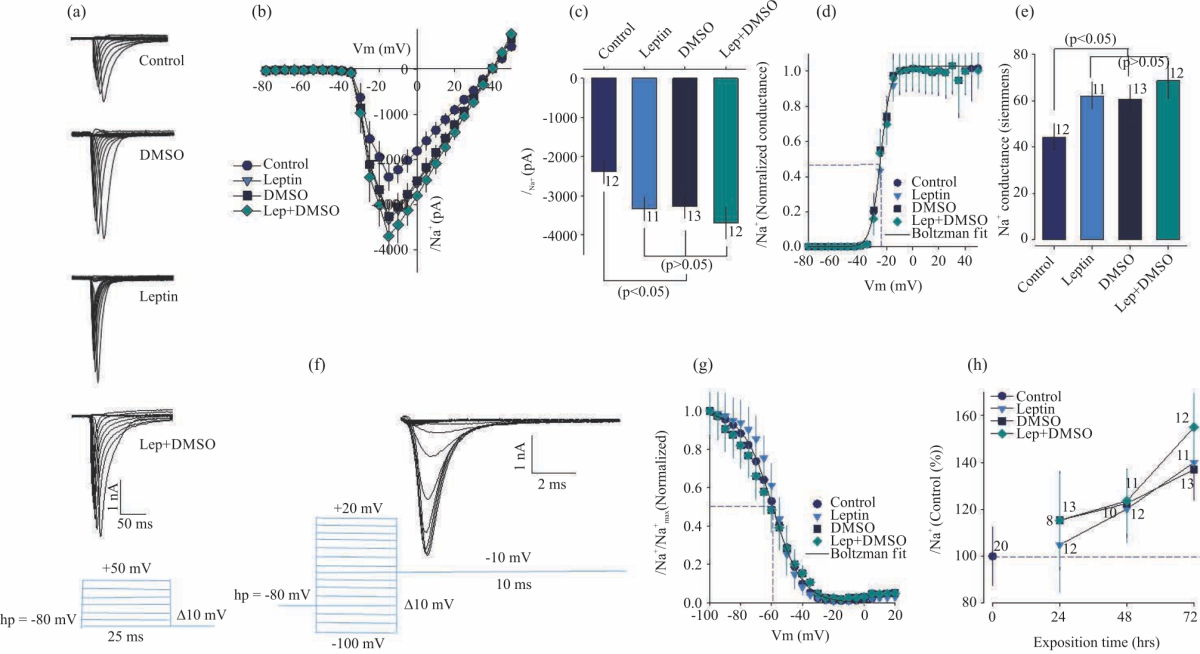 |
| Fig. 4(a-h): | Leptin enhances functional expression of the Na+ channel in neuroblastoma cell line N1E-115 (a) Trace Na+ family whole-cell currents (INa) evoked by depolarizing pulses from a holding potential (Vh) of -80mV to 50 mV, in 10 mV steps, in cells kept in culture for 72 hrs, in control or treatment conditions, (b) Average current-voltage (I-V) relationships obtained from control and treated cells with leptin, DMSO and leptin+DMSO, (c) Bar chart summarizing the effects of leptin or DMSO on INa, (d) Activation of Na+ currents, conductance-voltage relationships estimated from cells in (B), data were fitted with a Boltzmann equation, (e) Bar chart summarizing the effects of leptin or DMSO on Na+ conductance, (f) Trace family steady-state inactivation of INa+ evoked by a two-step pulse protocol: First, one conditioned pulse from a holding potential (Vh) of -80mV, from -100-20 mV in two 10 mV steps (10 ms duration); second, one test pulse to -20 mV (maximal INa+), with a duration of 10 ms, (g) Comparison of INa+ inactivation at steady state. Values in treated cells were normalized to the control values and (h) Time course dependence of stimulatory action of leptin or DMSO on INa+ current was converted to a percentage of its average value in control cells, *Significant differences (p<0.05) compared with control cells |
| Table 4: Fit values from the boltzmann equation of Na+ current kinetic properties in neuroblastoma N1E-115 cells treated with leptin or DMSO | ||||
| Kinetic properties | Experimental conditions | Max conductance (S) | Slope factor slope factor (k) pA/mV | Midpoint (v1/2) (mV) |
| Activation | Control | 44.27±5.83* | 3.19±0.44 | -21.53±2.44 |
| Leptin | 61.98±5.97 | 2.78±0.54 | -18.89±2.35 | |
| DMSO | 60.65±5.97 | 2.98±0.89 | -20.10±2.68 | |
| Leptin+DMSO | 68.62±7.87 | 3.01±0.24 | -19.65±2.31 | |
| Mean±EE | 58.88±6.41 | 2.99±0.52 | -20.04±2.44 | |
| Max current (nA) | Slope factor (k) pA/mV | Midpoint (v1/2) (mV) | ||
| Inactivation | Control | 2.84±0.37* | 9.54±1.27 | -59.20±2.34 |
| Leptin | 3.95±0.25 | 7.93±1.18 | -56.80±2.21 | |
| DMSO | 4.01±0.38 | 11.44±1.47 | -60.01±2.68 | |
| Leptin+DMSO | 4.21±0.51 | 8.24±1.18 | -57.53±2.21 | |
| Mean±EE | 3.75±0.37 | 9.28±1.27 | -58.38±2.36 | |
(*)Indicate significant differences (Tukey p<0.05) to the control group. DMSO: Dimethyl sulfoxide | ||||
Chronic treatment (72 hrs) with leptin or DMSO, increases sodium current (~1.8 folds) amplitude (Fig. 4b, c), without modifying the kinetic properties of activation (Fig. 4d). Leptin and DMSO increase sodium conductance (Fig. 4e), without modifying the kinetic properties of inactivation (Fig. 4f,g). Besides, the increase of sodium current observed is time-dependent (Fig. 4h). The Boltzmann equation fit values of all treatments are shown in Table 4. The mean values found in the present research for sodium channels activation kinetics were: Slope factor = 2.99±0.52 pA mV–1, Midpoint = -20.04±2.44 mV and Maximum conductance = 58.88±6.41 S. The mean values found for sodium channels inactivation kinetics were: Slope factor = 9.28±1.27 pA mV–1, Midpoint = -58.38±2.36 mV and Maximum current = 3.75±0.37 nA.
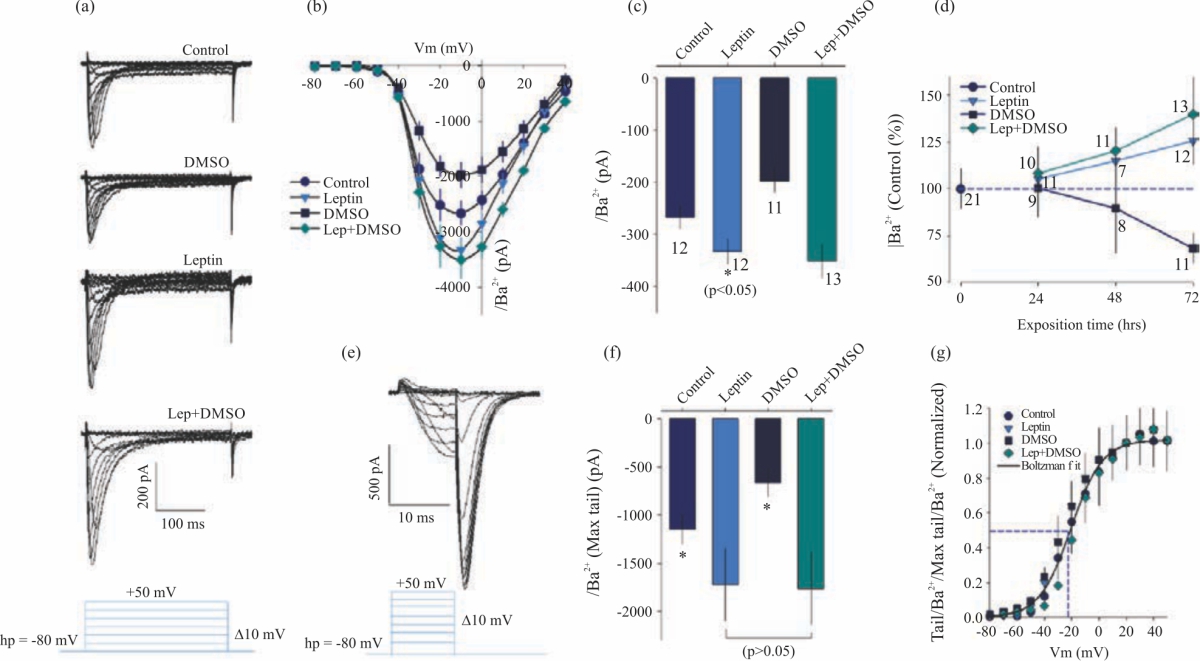 |
| Fig. 5(a-g): | Long-term treatment with leptin enhances T-Type Ca2+ channel activity in neuroblastoma cell line N1E-115. A (a) Trace family of voltage-activated Ba2+ currents (through Ca2+ channels) evoked by a depolarizing pulse test from a Vh of -80 mV, with a 10-mV interval in cell line N1E-115, kept in culture, in absence or presence of leptin or DMSO, (b) Average current-voltage (I–V) relationships obtained from control and treated cells, measured at the peak of each current trace, (c) Bar chart summarizing the effects of leptin or DMSO on IBa2+, (d) Time course dependence of stimulatory action of leptin or DMSO on IBa2+, the current was converted to a percentage of its average value in control cells, *Significant differences (p<0.05) compared with control cells, (e) Tail Ba2+ currents through Ca2+ channels evoked by 10-ms voltage steps from -80-+50 mV and were closed (deactivated) by the return to the Vh, (f) Bar chart summarizing the effects of Leptin or DMSO on Ba2+ maximal tail current and (g) Current-voltage relationship normalized to its maximal value tail current for T-Type currents, estimated from the tail current analysis. Data points were fitted with Boltzmann equation |
Leptin effects on ICa2+ current in neuroblastoma cell line N1E-115: Another aspect that was picked for analysis was whether the calcium current component was affected by chronic treatment with leptin or DMSO since an increase was observed in the inward currents. It has been reported that, in neuroblastoma N1E-115 line cells, the main component of calcium current is that which flows from Low Voltage-Activated (T-Type) Channels (LVA), the current component that flows from High Voltage-Activated Channels (HVA) is slight. With the use of Barium as a charge carrier, Barium current in N1E-115 cells was recorded under control conditions. N1E-115 cells show fast activation (<20 ms time to peak at -20 mV) and fast inactivation (<100 ms time decay) (Fig. 5a), which is consistent with a current that flows through Low Voltage-Activated (T-Type) Channels (LVA). Barium current begins to be observed at values close to -50 mV, reaching their peak between -20 and -10 mV (Fig. 5b). Leptin increases the barium current (~1.3 folds), DMSO decreases it (~1.3 folds) and leptin plus DMSO increases it in a similar amount to leptin (Fig. 5c), thus showing that leptin induces an increase in the barium current. The increase (leptin) and decrease (DMSO) of the barium current is time-dependent (Fig. 5d). The changes observed in the barium current, in cells treated with the drugs, do not modify the activation kinetic properties (Fig. 5e-g). The mean values found for calcium channels T-Type activation kinetics were: Slope factor = 11.39±2.70 pA mV–1, Midpoint = 22.95±2.83 mV and Maximum current = 1322±262 pA. The Boltzmann equation fit values of all treatments are shown in Table 5.
Molecular changes promoted by leptin in N1E-115 neuroblastoma cell line
Leptin effects on mRNA level expression of Nav, Cav, POMC and CART in neuroblastoma cell line N1E-115: The increases observed in the electrophysiological records of voltage-clamp in sodium and barium currents do not show any modification of the kinetic properties. This effect can be explained by an increase in the number of ion channels embedded in the plasma membrane of N1E-115 cells.
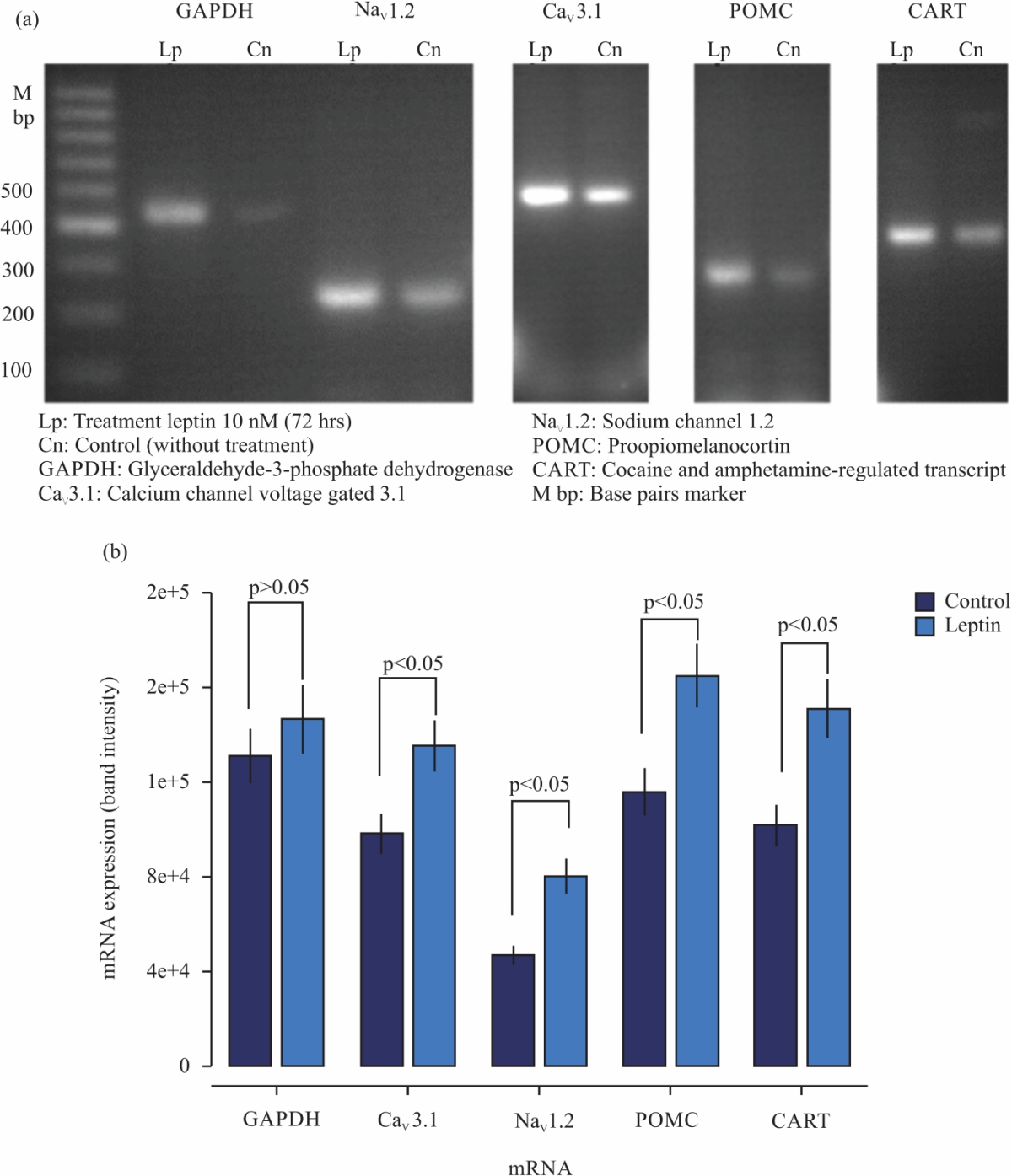 |
| Fig. 6(a-b): | Chronic effect of Leptin on mRNA level expression of Nav1.2, Cav3.1, POMC and CART in neuroblastoma cell line N1E-115 (a) Electrophoresis of Nav1.2, Cav3.1, POMC and CART mRNAs in control cells and cells treated with leptin 10 nM for 72 hrs and (b) Comparison of relative mRNA expression levels of Nav1.2 Cav3.1, POMC and CART normalized to GAPDH mRNA expression, “p” denotes significant differences (p<0.05) as compared with untreated cells |
| Table 5: Fit values from the Boltzmann equation of Ca2+ tail current kinetic properties in Neuroblastoma N1E-115 cells treated with leptin or DMSO | |||
| Experimental conditions | Max conductance (pA) | Slope factor (k) pA/mV–1 | Midpoint (v1/2) (mV) |
| Control | 1146±158* | 11.57±2.80 | 21.17±2.96 |
| Leptin | 1720±377 | 11.12±2.71 | 24.88±2.84 |
| DMSO | 663±139* | 12.44±2.61 | 24.91±2.73 |
| Leptin+DMSO | 1759±377 | 10.45±2.68 | 20.84±2.81 |
| Mean ± EE | 1322±262 | 11.39±2.70 | 22.95±2.83 |
(*) Indicate significant differences (Tukey p<0.05) to the control group. DMSO: Dimethyl sulfoxide | |||
To answer this question, molecular biology experiments (RT-PCR) were carried out. The mRNA level for the main subunit channel (Nav2.1 and Cav3.1) was determined in a semi-quantitative manner. Additionally, in this series of experiments, it was decided to analyze whether N1E-115 cells treated with leptin increased their mRNA level expression that codes for anorectic peptides (POMC and CART) since these cells produce and secrete these energy-balancing peptides. The data of Fig. 6, shows RT-PCR molecular biology analyses of N1E-115 cells treated with leptin [10 nM] for 72 hrs. The level of intensity in the leptin-treated bands of the different oligonucleotides analyzed shows a clear increase in mRNA level expression (Fig. 6a). With the aid of image intensity analysis software, the levels of expression of the mRNA encoded for the different peptides analyzed were semi-quantified (Fig. 6b). Leptin induces an increase in mRNA expression that codes for the main subunits of sodium (Nav1.2) and Calcium ion channels (Cav3.1), in N1E-115 cells, Besides, it is observed that leptin promotes an increase of level expression in the mRNA that encodes for POMC and CART peptides, thus showing that leptin regulates anorexigenic peptides in N1E-115 neuroblastoma cells.
DISCUSSION
The present study reports an in vitro model of the trophic (ion currents) and differentiation (neurite presence, dendrites and axons) effects of chronic treatment with leptin on “ARC-like” N1E-115 cells. Previously, N1E-115 has been used as an in vitro model of differentiated neuron cells18,20. The results of morphological analysis by light microscopy demonstrate for the first time that leptin promotes significant changes in its morphology and neurite formation of the N1E-115 cells, same results can be seen an in vivo models, in which energy deficiencies in utero life have been analyzed on changes in the arcuate nucleus axons pathways6,7,31. Oh et al.20, have shown that different signaling proteins, including GTP-binding /Ras-related proteins, growth factors, calcium-binding proteins and phosphatase-related proteins are involved in DMSO-differentiated N1E-115 cells. Avraham et al.32, have reported that leptin can induce neurogenesis and angiogenesis in Sabra mice who underwent photo-thrombotic stroke and increase of pAMPK concentrations, a potent AMP kinase inhibitor that is central to cell survival. In glial cells such as those found in the hypothalamus, chronic administration of leptin induce astrocyte structural protein expressions, with this, astrocyte morphology can be modified, these effects modulate synaptic inputs in the hypothalamus33. Different authors have shown the effects of leptin on the proliferation and differentiation of different cells, including non-neuronal cells, in the central nervous system34, mesenchymal progenitor35, chondrogenic progenitor36, marrow stromal37, preadipocytes38 and others. In the present study, the morphological differentiation promoted by leptin was studied without using any activation or inactivation signaling pathway drug. To clarify this event, molecular biology and signaling pathways experiments must be carried out to elucidate the genes that are expressed and signaling pathways involved in promoting proliferation and differentiation in in vitro models such as N1E-115 cells.
The neuronal electrophysiological differentiation is characterized by the ability of the cell to generate repetitive discharges, either spontaneously or in response to stimulation. In leptin-differentiated N1E-115 cells, an increase in the amplitude of sodium ion current is observed as the cell is depolarized. The increase in the amplitude of the sodium current did not modify the kinetic properties of the sodium channel, similar results have been shown in other cells treated with leptin as chromaffin cells39, gonadrotropes40, pancreatic β-cells41 and others. In this sense, additionally, to stimulating morphological changes in N1E-115 cells, leptin can increase the up-regulation of Na+ channels present in the plasma membrane of leptin-differentiated N1E-115 cells. Here, we show that the chronic treatment (72 hrs) with leptin specifically increases the functional expression of voltage-gated Na+ channels, which are responsible for the spiking phase of action potentials, our results support the idea that leptin can modulate the electrical activity in differentiated N1E-115 cells.
Non-differentiated N1E-115 neuroblastoma cells show two components of calcium current: A rapidly activating and rapidly inactivating current carried by LVA T-type Ca2+ channels and a slowly activating and non-inactivating current carried by HVA L-type Ca2+ channels42,43, with a predominance of calcium current carried by T-type Ca2+ channels44. In this series of experiments, Ba2+ [10nM] was used as a charge carrier to examine Ba2+ current through Ca2+ channels. DMSO-differentiated N1E-115 cells show a reduction in the Ba2+ current, while leptin-differentiated N1E-115 cells show increases in Ba2+ current. The rapidly inactivating component is active in the voltage range -60 mV<V<+60 mV and is maximal at ~-20 mV, its time constant is <100 ms43 and the rapidly inactivating component is essentially complete within 0.5 sec. When analyzing the kinetic activation properties using current tail analysis, no changes were observed in the kinetic activation properties of the T-type Ca2+ channel. The discrepancy in the Ba2+ current results may be because the two drugs (DMSO or Leptin) have been reported to use different signaling pathways20. Increments in Ca2+ currents have been reported in other cells treated with leptin, such as chromaffin cells39, pituitary cells40, pancreatic β-cells41 and others. In this sense, leptin can increase the up-regulation of Ca2+ ion channels present in the plasma membrane of leptin-differentiated N1E-115 cells and profoundly affect cellular excitability by increasing the amplitude of the T-type currents. The presence of Na+ channel 1.2 (Nav1.2) has been reported in many areas of the CNS, including the hypothalamus44, furthermore to being present in N1E-115 cells28. The results show an increase in Na+ current amplitude due to an up-regulation of Na+ channels in the plasma membrane of leptin-differentiated N1E-115 cells, this effect is a consequence of signaling pathway stimulation aimed at activating the gene that codes sodium channel α-subunits, reflected in increased expression levels of mRNA sodium channel α-subunits (Nav1.2). N1E-115 cells express the Ca2+ channel isoform Cav3.1, which plays a crucial role in determining neuronal excitability29. It is recognized that Ca2+ channels modulate the function of different neuronal types by influencing synaptic transmission and neuronal excitability42. Neuronal T-type channels have been shown to promote burst firing and low-amplitude intrinsic neuronal oscillations as well as Ca2+ entry and amplification of dendritic synaptic signals. Last, leptin can regulate levels of CART/POMC or AGRP/NPY gene expression11,16. Non-differentiated N1E-115 cells contain mRNA for POMC, CART, Ob-Rb21. Chronic treatment with leptin as well as stimulating cell differentiation in N1E-115 cells, promotes an increase in the levels of expression of the mRNA that encodes POMC and CART as mentioned in results and thus, show an in vitro model of cell differentiation on “ARC-like” neurons.
The clinical implications of the results can be compared with those observed in utero development studies, where the reduced axonal pathways of ARC results from leptin deficiency may contribute to appetite dysregulation, furthermore, there may be a closer impact on the formation of a sufficient population of neurons within the ARC. During development and differentiation as can be observed in in vitro or in vivo models, neuro-progenitor cell undergoes extensive self-renewal and then generate neurons and astrocytes, an altered hormone or neurotrophic environment may alter the critical neurodevelopmental signaling of ARC.
CONCLUSION
Leptin chronic treatment promotes neurite expression (Axons and Dendrites), an increment in electrical excitability by increasing the sodium and calcium ion currents due to Up-regulation in mRNA expression that codes for the main subunits of sodium (Nav1.2) and Calcium (Cav3.1) ion channels, besides increasing the mRNA level expression of anorexic peptides Pro-Opiomelanocortin (POMC) and cocaine and amphetamine-regulated transcript (CART) in “ARC-like” neuroblastoma N1E-115.
SIGNIFICANCE STATEMENT
This study discovers the possible effect of chronic leptin treatment on differentiation and expression of Na+ and Ca2+ channels in the N1E-115 cell line used as a model of arcuate nucleus cells, which can be beneficial for analyzing the neuronal differentiation and axonal pathways in the Hypothalamus. This study will help the researcher to uncover signaling pathways using hormones of energy balance on neuronal development that many researchers were not able to explore. Thus, a new theory on the effects of orexigenic and anorexigenic peptides may be arrived at.
ACKNOWLEDGMENTS
RIVR was supported by fellowships from Conacyt (Mexico). The authors would like to thank Dr. Ricardo Felix (CINVESTAV) for the kindly donation of N1E-115 cells. This work was supported by the Academic Staff Biotechnology and Reproduction (UVE-CA-252)
REFERENCES
- Llinás, R.R., 2014. Intrinsic electrical properties of mammalian neurons and CNS function: A historical perspective. Front. Cell. Neurosci., Vol. 8.
CrossRefDirect Link - Nogueiras, R., M.H. Tschöp and J.M. Zigman, 2008. Central nervous system regulation of energy metabolism. Ann. New York Acad. Sci., 1126: 14-19.
CrossRefDirect Link - Zhang, Y., R. Proenca, M. Maffei, M. Barone, L. Leopold and J.M. Friedman, 1994. Positional cloning of the mouse obese gene and its human homologue. Nature, 372: 425-432.
CrossRefDirect Link - Breslow, M.J., K. Min-Lee, D.R. Brown, V.P. Chacko, D. Palmer and D.E. Berkowitz, 1999. Effect of leptin deficiency on metabolic rate inob/obmice. Am. J. Physiol. Endocrinol. Metab., 276: E443-E449.
CrossRefDirect Link - Cohen, P., C. Zhao, X. Cai, J.M. Montez and S.C. Rohani et al., 2001. Selective deletion of leptin receptor in neurons leads to obesity. J. Clin. Invest., 108: 1113-1121.
CrossRefDirect Link - Desai, M., T. Li and M.G. Ross, 2011. Fetal hypothalamic neuroprogenitor cell culture: preferential differentiation paths induced by leptin and insulin. Endocrinology, 152: 3192-3201.
CrossRefDirect Link - Desai, M., T. Li and M.G. Ross, 2011. Hypothalamic neurosphere progenitor cells in low birth-weight rat newborns: Neurotrophic effects of leptin and insulin. Brain Res., 1378: 29-42.
CrossRefDirect Link - Heijboer, A.C., H. Pijl, A.M. Van den Hoek, L.M. Havekes, J.A. Romijn and E.P. Corssmit, 2006. Gut-brain axis: Regulation of glucose metabolism. J. Neuroendocrinol., 18: 883-894.
CrossRefDirect Link - Broberger, C., J. Johansen, C. Johansson, M. Schalling and T. Hokfelt, 1998. The neuropeptide Y/agouti gene-related protein (AGRP) brain circuitry in normal, anorectic, and monosodium glutamate-treated mice Proc. Nat. Acad. Sci., U.S. Am., 95: 15043-15048.
CrossRefDirect Link - Elias, C.F., C. Lee, J. Kelly, C. Aschkenasi and R.S. Ahima et al., 1998. Leptin activates hypothalamic CART neurons projecting to the spinal cord. Neuron, 21: 1375-1385.
CrossRefDirect Link - Bouret, S.G., S.J. Draper and R.B. Simerly, 2004. Trophic action of leptin on hypothalamic neurons that regulate feeding. Science, 304: 108-110.
CrossRefDirect Link - Desai, M., D. Gayle, J. Babu and M.G. Ross, 2007. The timing of nutrient restriction during rat pregnancy/lactation alters metabolic syndrome phenotype. Am. J. Obstetrics Gynecol., 196: 555.e1-555.e7.
CrossRefDirect Link - Lang, B.T., Y. Yan, R.J. Dempsey and R. Vemuganti, 2009. Impaired neurogenesis in adult type-2 diabetic rats. Brain Res., 1258: 25-33.
CrossRefDirect Link - Marks, J.L., D.J. Porte, W.L. Stahl and D.G. Baskin, 1990. Localization of insulin receptor mRNA in rat brain by in situ hybridization. Endocrinology, 127: 3234-3236.
CrossRefDirect Link - Cheung, C.C, D.K. Clifton and R.A. Steiner, 1997. Proopiomelanocortin neurons are direct targets for leptin in the hypothalamus. Endocrinology, 138: 4489-4492.
CrossRefDirect Link - Grimbly, C., B. Martin, E. Karpinski and S. Harvey, 2009. Growth hormone production and action in N1E-115 neuroblastoma cells. J. Mol. Neurosci., 39: 117-124.
CrossRefDirect Link - Kimhi, Y., C. Palfrey, I. Spector, Y. Barak and U.Z. Littauer, 1976. Maturation of neuroblastoma cells in the presence of dimethylsulfoxide. Proc. Nat. Acad. Sci., U.S. Am., 73: 462-466.
CrossRefDirect Link - Marler, K.J., R. Kozma, S. Ahmed, J.M. Dong, C. Hall and L. Lim, 2005. Outgrowth of neurites from NIE-115 neuroblastoma cells is prevented on repulsive substrates through the action of PAK. Mol. Cell. Biol., 25: 5226-5241.
CrossRefDirect Link - Oh, J.E., K.R. Karlmark, J.H. Shin, A. Pollak, A. Freilinger, M. Hengstschläger and G. Lubec, 2005. Differentiation of neuroblastoma cell line N1E-115 involves several signaling cascades. Neurochem. Res., 30: 333-348.
CrossRefDirect Link - Roth, J.D., D.K. Yee, L.R. Kisley and S.J. Fluharty, 2002. Modeling the pathways of energy balance using the N1E-115 murine neuroblastoma cell line. Mol. Brain Res., 103: 146-150.
CrossRefDirect Link - Dominguez, B., R. Felix and E. Monjaraz, 2009. Upregulation of voltage-gated Na+ channels by long-term activation of the ghrelin-growth hormone secretagogue receptor in clonal GC somatotropes. Am. J. Physiol.-Endocrinol. Metab., 296: E1148-E1156.
CrossRefDirect Link - Avila, G., A. Sandoval and R. Felix, 2004. Intramembrane charge movement associated with endogenous K+ channel activity in HEK-293 cells. Cell. Mol. Neurobiol., 24: 317-330.
CrossRefDirect Link - Dubois, J.M., G. Ouanounou and B. Rouzaire-Dubois, 2009. The boltzmann equation in molecular biology. Prog. Biophys. Mol. Biol., 99: 87-93.
CrossRefDirect Link - Matteson, D.R. and C.M. Armstrong, 1986. Properties of two types of calcium channels in clonal pituitary cells. J. Gen. Physiol., 87: 161-182.
CrossRefDirect Link - Meza, U., G. Avila, R. Felix, J.C. Gomora and G. Cota, 1994. Long-term regulation of calcium channels in clonal pituitary cells by epidermal growth factor, insulin and glucocorticoids. J. Gen. Physiol., 104: 1019-1038.
CrossRefDirect Link - Dominguez, B., T. Avila, J. Flores-Hernandez, G. Lopez-Lopez, H. Martinez-Rodriguez, R. Felix and E. Monjaraz, 2008. Up-regulation of high voltage-activated Ca2+ channels in gc somatotropes after long-term exposure to ghrelin and growth hormone releasing peptide-6. Cell. Mol. Neurobiol., 28: 819-831.
CrossRefDirect Link - Baroni, D. and O. Moran, 2011. Molecular differential expression of voltage-gated sodium channel α and β subunit mRNAs in five different mammalian cell lines. J. Bioenerg. Biomembr., 43: 729-738.
CrossRefDirect Link - Calderón-Rivera, A., A. Sandoval, R. González-Ramírez, C. González-Billault and R. Felix, 2015. Regulation of neuronal cav3.1 channels by cyclin-dependent kinase 5 (Cdk5). PLoS ONE, Vol. 10.
CrossRefDirect Link - Church, D.M., V.A. Schneider, T. Graves, K. Auger and F. Cunningham et al., 2011. Modernizing reference genome assemblies. PLoS Biol., Vol. 9.
CrossRefDirect Link - Valassi, E., M. Scacchi and F. Cavagnini, 2008. Neuroendocrine control of food intake. Nut. Metab. Cardiovasc. Dis., 18: 158-168.
CrossRefDirect Link - Avraham, Y., N. Davidi, V. Lassri, L. Vorobiev and M. Kabesa et al., 2011. Leptin induces neuroprotection neurogenesis and angiogenesis after stroke. Curr. Neurovasc. Res., 8: 313-322.
CrossRefDirect Link - García-Cáceres, C., E. Fuente-Martín, E. Burgos-Ramos, M. Granado and L.M. Frago, 2011. Differential acute and chronic effects of leptin on hypothalamic astrocyte morphology and synaptic protein levels. Endocrinology, 152: 1809-1818.
CrossRefDirect Link - Fujita, Y. and T. Yamashita, 2019. The effects of leptin on glial cells in neurological diseases. Front. Neurosci., Vol. 13.
CrossRefDirect Link - Scheller, E.L., J. Song, M.I. Dishowitz, F.N. Soki, K.D. Hankenson and P.H. Krebsbach, 2010. Leptin functions peripherally to regulate differentiation of mesenchymal progenitor cells. Stem cells, 28: 1071-1080.
CrossRefDirect Link - Zhao, X., Y. Dong, J. Zhang, D. Li and G. Hu et al., 2016. Leptin changes differentiation fate and induces senescence in chondrogenic progenitor cells. Cell Death Dis., Vol. 7.
CrossRefDirect Link - Thomas, T., F. Gori, S. Khosla, M.D. Jensen, B. Burguera and B.L. Riggs, 1999. Leptin acts on human marrow stromal cells to enhance differentiation to osteoblasts and to inhibit differentiation to adipocytes. Endocrinology, 140: 1630-1638.
CrossRefPubMedDirect Link - Patel, N.G., J.C. Holder, S.A. Smith, S. Kumar and M.C. Eggo, 2003. Differential regulation of lipogenesis and leptin production by independent signaling pathways and rosiglitazone during human adipocyte differentiation. Diabetes, 52: 43-50.
CrossRefDirect Link - Takekoshi, K., M. Motooka, K. Isobe, F. Nomura, T. Manmoku, K. Ishii and T. Nakai, 1999. Leptin directly stimulates catecholamine secretion and synthesis in cultured porcine adrenal medullary chromaffin cells. Biochem. Biophys. Res. Commun., 261: 426-431.
CrossRefDirect Link - Domínguez-Mancera, B., M. Barrientos-Morales, P. Cervantes-Acosta, A. Hernández-Beltrán and A. González-Ramírez et al., 2017. Leptin regulation of inward membrane currents, electrical activity and lh release in isolated bovine gonadotropes. Biochem. Biophys. Res. Commun., 491: 53-58.
CrossRefDirect Link - Domínguez-Mancera B., A. Rodríguez-Andrade, M. Barrientos-Morales, P. Cervantes-Acosta and A. Hernández-Beltrán, 2019. Ghrelin and Leptin increase voltage-gated Na+ and Ca2+ currents in the RIN-m5f insulin-producing cell line. Indian J. Biochem. Biophys., 56: 193-204.
Direct Link - Perez-Reyes, E., 2003. Molecular physiology of low-voltage-activated t-type calcium channels. Physiol. Rev., 83: 117-161.
CrossRefDirect Link - Bolsover, S.R., 1986. Two components of voltage-dependent calcium influx in mouse neuroblastoma cells. Measurement with arsenazo III. J. Gen. Physiol., 88: 149-165.
CrossRefDirect Link - Kwong, K. and M.J. Carr, 2015. Voltage-gated sodium channels. Curr. Opin. Pharmacol., 22: 131-139.
CrossRefDirect Link