
Hayat Ali Alafari
Department of Biology, Faculty of Science, Princess Nourah Bint Abdulrahman University, Riyadh, Saudi Arabia
Magda Elsayed Abd-Elgawad
Department of Biology, Faculty of Science, Princess Nourah Bint Abdulrahman University, Riyadh, Saudi Arabia
LiveDNA: 20.32558
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 3 | Page No.: 310-318







ABSTRACT
Background and Objective: Heat stress is known as a raise of 5°C or more than the optimal temperature. In this study, we analyzed the effect of heat stress on protein content, protein electrophoretic pattern and Superoxide Dismutase (SOD) profile in three populations of Tetraena propinqua subspecies migahidii. Materials and Methods: Populations of Tetraena propinqua ssp. migahidii were studied. The seeds were subjected to 25 (control), 30, 35 and 40°C for 4, 24 and 48 hrs and 10 days. Results: Heat stress (35 and 40°C) elicited total soluble protein in populations 1 and 2 however reduced in population 3 with increasing exposure time to 10 days. New polypeptides of 23 KD at 4 hrs in population 3 below 35°C and population 2 below 40°C and 28 KD at 48 hrs in population one below 30°C however 20 KD altogether populations below 40°C. The expression of most polypeptides diminished for 4 hrs however induced for 24, 48 hrs and 10 days with increase heat temperature to 40°C relative to their expression among the management seedlings. SOD1 and SOD2 have detected altogether most of the genotypes, however, heat stress (35, 40°C) induced the expression of SOD2 and SOD1 and was altogether genotyped for 10 days as compared with the control. Conclusion: The heat stress caused protein degradation and conjointly induced expression of new synthesized HSPs throughout heat acclimatization may be related to heat injury and the improved thermotolerance in early hours of germination and additional studies are required for its protein identification.
PDF Abstract XML References Citation
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Hayat Ali Alafari and Magda Elsayed Abd-Elgawad, 2021. Heat-Induced Protein and Superoxide Dismutase Changes in Wild Tetraena propinqua ssp. Migahidii Seedlings. Pakistan Journal of Biological Sciences, 24: 310-318.
DOI: 10.3923/pjbs.2021.310.318
URL: https://scialert.net/abstract/?doi=pjbs.2021.310.318
DOI: 10.3923/pjbs.2021.310.318
URL: https://scialert.net/abstract/?doi=pjbs.2021.310.318
INTRODUCTION
A variety of external stresses like cold, heat, water deficit or drought, flooding, high salinity and strong winds are the most harmful factors regarding the growth, development and survival of sessile plants. The external stresses can purpose for the expulsion of Reactive Oxygen Species (ROS) including hydrogen peroxide (H2O2), superoxide anion (·O–2), singlet oxygen (1O2) and hydroxyl radical (·OH)1-3. Heat stress is known as a raise of 5°C or more than the optimal temperature4. Heat Stress (HS) gives rise to an excessive increase in membrane fluidity, a confusion of protein function and turnover and metabolic imbalances in plants5. To abolish the detrimental effects of heat stress, plants have appointed multiple stress-tolerant planning’s by changing the gene expression, protein synthesis and post-translational modification, which participate in the repetition of cellular homeostasis for plant survival under high temperature6. The plant cells interact with heat stress by stimulating the transcription of genes encoding heat shock proteins (Hsp), which are implicated in preventing or repairing the damage caused by elevated temperature and thus give increased thermotolerance7.
Heat-shock proteins (Hsps) task as molecular chaperones that support protein folding and inhibit irreversible protein accumulation to preserve cellular homeostasis below each optimum and adverse developmental conditions8. Instituted on their approximate molecular weights, five major families of Hsps are known: Hsp100, Hsp90, Hsp70, Hsp60 and the small Hsp (sHsp) families.
Heat stress has been reported to stimulate the overproduction of Reactive Oxygen Species (ROS)9 but plants may also reactivate antioxidant enzymes, such as Superoxide Dismutase (SOD) and ascorbate peroxidase (APX), to remove ROS or to mitigate the impact of ROS10. SOD, a member of the metalloenzyme family, induces the disharmony of O2– ion into hydrogen peroxide (H2O2) and oxygen (O2) molecules. SOD has been qualified to be implicated in several abiotic stress-related responses like heat, drought, salt, low-temperature conditions11-13.
Tetraena was treated as a taxon genus and its sole species, it’s classified by Beier14 as a genus of Zygophylloideae. Currently, Tetraena includes concerning 40 species with a distribution extending from the Canary Islands within the west, South Africa within the South to China within the East, of that 8 species are found in Saudi Arabia15. However, T. propinqua differs from alternative Tetraena species in some morphological characteristics like leaves 2-foliolate, the color of flower and fruit form. The conservation status of T. propinqua ssp. migahidii seems to be distributed in some localities within the north region and western central of Saudi Arabia. It’s habitats sandy salt habitats and gravels desert. T. propinqua ssp. migahidii, is distinguished by leaves with apex rounded, white-creamy flowers and capsules obconical16.
Therefore, the aims of this study were planned to look at the result of heat stress on the content of total soluble proteins, SDS-PAGE and superoxide dismutase antioxidant enzyme of T. propinqua ssp. migahidii populations.
MATERIALS AND METHODS
Samples collection: Seeds of wild T. propinqua ssp. migahidii were collected from May-July, 2018 from plants growing in central Saudi Arabia (Fig. 1). Three samples of T. propinqua ssp. migahidii was collected from population 1, Riyadh-Al Kharj road (Al Kharj expressway), population 2, Riyadh-Al Qassim road (King Salman road 3306) and population 3, Al Thumama road (Al Narjas) (Fig. 1). The study was carried out at Department of genetic origins, Biochemistry Lab, Egypt from March-December, 2019) Seeds were collected from several separate plants selected randomly throughout the whole region, used for various germination experiments.
Seeds germination: Germination of T. propinqua ssp. migahidii in response to heat was investigated in genetics lab by placing 50 seeds in a 9-cm-diameter Petri dish containing two layers of Whatman No. 1 filter papers, (Whatman, China) moistened with 5 mL of distilled water and 3 mL added after 3 days’ interval. The Petri dishes were incubated at 25, 30, 35 and 40°C in complete darkness. The samples were collected at 4, 24, 48 hrs and10 days. The experiment was replicated three times.
Extraction: Seed materials (50 seeds) were homogenized in cold 100 mM phosphate buffer pH 7.8 containing 0.1 EDTA. Homogenates were centrifuged at 12000 rpm for 20 min to remove plant debris. Supernatants were used as samples for measuring soluble protein content, SDS-PAGE of soluble proteins and Superoxide dismutase (SOD) electrophoresis.
Soluble protein quantification: Extracts of different samples were tested for their protein concentrations according to the microassay method of Bradford 17. Duplicate samples in a total volume of 100 μL were pipetted into 1.5 mL polyethylene microfuge tubes (BrandTM Polypropylene Microcentrifuge Tubes).
 |
| Fig. 1: | Distribution of T. propinqua ssp. migahidii populations in different natural habitats of central Saudi Arabia Three populations were, Population 1: Riyadh-Al Kharj road (Al Kharj expressway), Population 2: Riyadh-Al Qassim road (King Salman road 3306), Population 3: Al Thumama road (Al Narjas) Source: https://www.express.co.uk/news/world/1032999/Saudi-Arabia-map-where-is-Saudi-Arabia-what-countries-surround- Saudi-Arabia |
Protein reagent (1 mL) was added to each tube and mixed thoroughly and gently. The assay reagent is made by dissolving 100 mg of Coomassie Blue G250 in 50 mL of 95% ethanol. The solution is then mixed with 100 mL of 85% phosphoric acid and made up to 1 L with distilled water. The absorbance of each sample between 2 and 60 min after the addition of the protein reagent was measured at 280 nm using Biorad® spectrophotometer (Bio-Rad Laboratories, India). Bovine serum albumin was used as a standard.
SDS-PAGE of soluble proteins: Samples extracts were mixed with sample loading buffer in suitable amounts to obtain equal protein concentration for all samples. The loading buffer was containing Tris- HCl (0.5M pH 6.8), EDTA (0.25 M) SDS (5%), Glycerol (10%), 2 mg bromophenol blue, (7.15M) b-mercaptoethanol and ddH2O up to 10 mL. Samples were loaded into the gel on a vertical slab using a Biometra apparatus according to the method of Laemmli18. The run buffer was added to the upper tank just before running. This gel was run at 80 volts for a quarter of an h, then the voltage was raised to 120 volts until the samples reach one inch from the bottom of the gel. The gel was removed from the apparatus gently and placed in a plastic tank, then covered with 100 mL of fixing solution overnight. Then, the gel was stained with Coomassie brilliant blue (CBB-G250) stain solution proposed by Kang et al.19 and gently agitated for 2 hrs. The prestained protein ladder used as a marker with Molecular Weights (MW) ranged between 10-245 KD (GangNam-STAINTM). Stained gel was photographed and analyzed separately using GelAnalyzer® software.
SOD native PAGE (Superoxide dismutase on native polyacrylamide gels): The used protocol was described by Chen and Pan20 with modification done by Weydert and Cullen21 as follows: Sample extracts were mixed with sample loading buffer in suitable amounts to obtain equal protein concentration for all samples. To prepare 2 mL of sample loading buffer mix 2 mL of sucrose (40%) and 20 μL of 5% bromophenol blue solution (wt/vol). Samples were loaded into the gel which containing two separating mini-gels of separating gel 12% acrylamide and stacking gel 6% acrylamide, while in the pre-electrophoresis buffer and then the electrophoresis system was operated at 80 volts. After 15 min, the run was stopped and the buffer was replaced with the electrophoresis running buffer to continue run at 120 volts. As the bromophenol blue stain reached the end of the gel, the run was stopped. The gel was pulled out gently from the glass plates and washed with distilled water. The washed gel was submerged in native SOD staining solution, incubated for 20 min in dark with shaking and then rinsed gently with ddH2O twice. Enough ddH2O was added to cover the gel and placed under a fluorescent light and on a light box. The gel begins to turn blue/purple and clear bands appear gradually. Achromatic bands indicate the presence of SOD. The gel was imaged using Biorad® GelDoc-XR imaging systems (Bio-Rad Laboratories, Segrate (Milan) Italy).
Statistical analysis: All values reported in experiments for protein content measurements are mean of three replicates. Data were analyzed with Past3 and the Excel software was used to construct diagrams.
RESULTS
Effect of heat stress on the soluble protein content of germinated seeds: As shown in (Fig. 2a, d), a major increase within the total soluble protein content bit by bit at 4 hrs of germination beneath heat temperature 35°C in population 1 and 3 (131.6 and 132.9 μg/10 μL), however, it decreased in population 2 (109.6 μg/10 μL) then decreased beneath 40°C altogether populations (117.8, 112.1 and 124 μg/10 μL) as compared with the conventional temperature (25°C during this study) (128.9, 121.2 and 115.1 μg/10 μL), severally (Fig. 2a). At 24 hrs of germination, the whole soluble protein decreased bit by bit in population one (110.6 μg/10 μL) beneath 40°C as compared with the conventional temperature (127.1 μg/10 μL), however, increased beneath heat temperature 35°C in population 3 (124 μg/10 μL) and beneath 40°C in population two and three (121.6 and 123.5 μg/10 μL) than normal temperature (113.1 and 115.1 and μg/10 μL), severally (Fig. 2b).
When 48 hrs of germination, it increased altogether populations once the temperature was increased to 40°C (132.3, 128.4 and 135.6 μg/10 μL) as compared to manage (124.7, 113.1 and 116.1 μg/10 μL), severally (Fig. 2c). When 10 days, it decreased once the temperature was increased to 40°C in population two and three (107.2 and 109.1 μg/10 μL), otherwise increased in population one (137 μg/10 μL) as compared to manage (113.1, 139.2, 133.6 μg/10 μL), severally (Fig. 2d).
Effects of heat stress on SDS-PAGE of soluble protein profile: For SDS-PAGE, proteins were extracted from germinating seeds of heat-treated and heat-untreated seedlings. Comparison of the protein profiles between control and those treated with completely different heat treatments using SDS-PAGE showed that heat treatment-induced solely a couple of changes within the pattern of proteins.
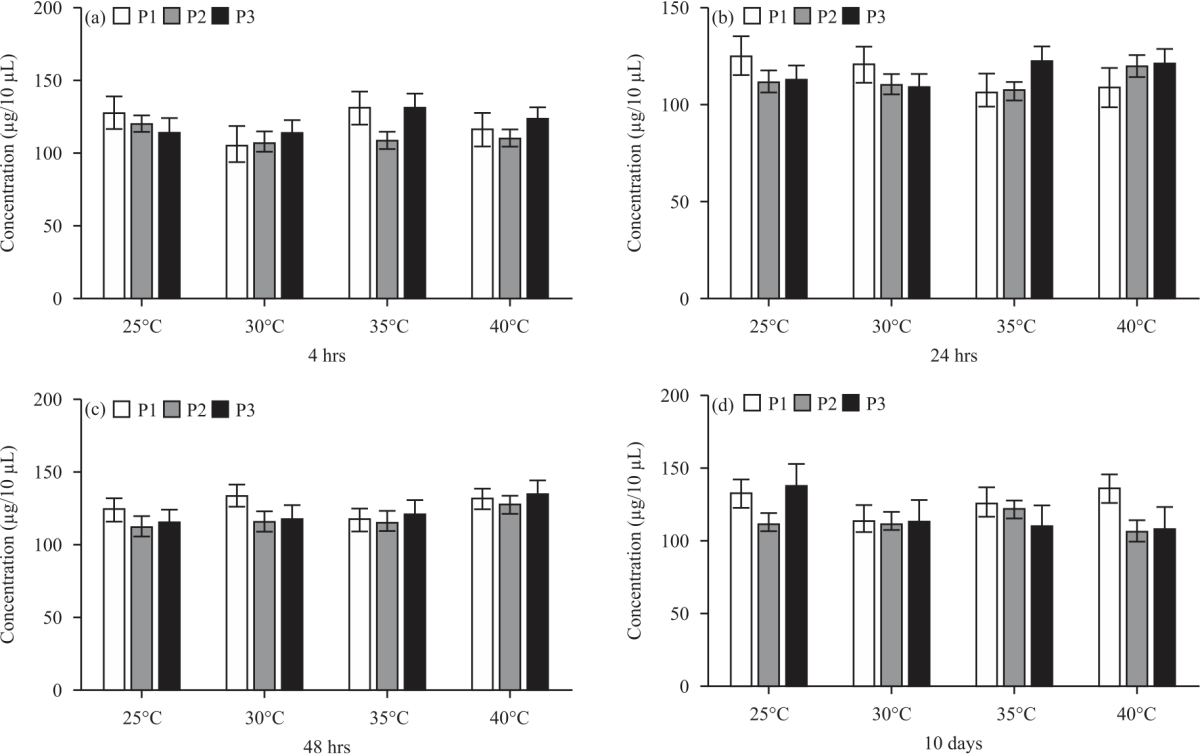 |
| Fig. 2(a-d): | Soluble protein content in seedlings of three populations of Tetraena propinqua under heat stress after exposure for (a) 4 hrs, (b) 24 hrs, (c) 48 hrs and (d) 10 days at each temperature (25, 30, 35, 40°C) |
 |
| Fig. 3(a-d): | SDS-PAGE of Soluble Proteins in seedlings of three populations of Tetraena propinqua under heat stress after exposure for (a) 4 hrs, (b) 24 hrs, (c) 48 hrs and (d) 10 days at each temperature 25°C (1, 2, 3); 30°C (4, 5, 6); 35°C (7, 8, 9); 40°C (10, 11, 12). Population 1: (1, 4, 7, 10), Population 2: (2, 5, 8, 11) and Population 3: (3, 6, 9, 12) |
At 4 hrs of germination, results showed that the looks of nine polypeptides of 25, 30, 37, 39, 40, 56, 58, 68, 83 and 96 KD altogether populations underneath untreated conditions however that of 39 KD absent in population one. Some polypeptides disappeared as 39 KD underneath (30-40°C) in population 2, underneath 40°C in population 3 and 25 KD in population 1 underneath 30°C, two severally, however, the peptide of 23 KD was newly synthesized underneath heat temperature 35 and 40°C in population 3 and 2, respectively. The intensity of bands minimized with increase heat temperature to 40°C as compared to the management (Fig. 3a). At 24 hrs of germination, Fig. 3b showed that the synthesis of 3 polypeptides of 26 KD ascertained all told populations under-treated and untreated heat temperature, 53 KD all told populations beneath untreated seedlings, in population one and two beneath (30 and 40°C) severally and 23 KD in population three beneath (25 and 35°C), in population one beneath (30 and 35°C) and all told populations beneath 40°C. Otherwise, some polypeptides of 83 and 68 KD disappeared all told populations beneath heat temperature 35°C. The band intensity increased with increase heat temperature to 40°C as compared to their intensities within the management plants. At 48 hrs of germination, polypeptides of 83 and 68 KD disappeared solely beneath normal temperature in population two and three and polypeptides of 40 KD disappeared once heat temperature increase than normal temperature, however, polypeptides of 39 KD observed all told populations once heat temperature increase than normal temperature. Some polypeptides recently synthesized as 28 KD in population one beneath 30°C and 20 KD all told populations beneath 40°C. The band intensity increased with increase heat temperature relative to their reference quantities within the management seedlings (Fig. 3c). Once ten days of germination, some polypeptides disappeared as 40 KD beneath untreated and treated heat temperature 40°C and 30 KD beneath untreated and treated expect in population one beneath 40°C, however that of 2 polypeptides of 25 KD and 23 KD appeared beneath 40, 30 and 35°C severally. The band intensity increased with an increased in heat temperature as compared to the management seedlings (Fig. 3d).
Effects of heat stress on superoxide dismutase antioxidant enzyme: All 3 populations had SOD1, SOD2 or both of them isozymes below heat-treated and heat-untreated seedlings (Fig. 3a, d). The intensity of SOD1 with (Rf 0.27) shrunken bit by bit at 4 hrs once the temperature was increased bit by bit to 40°C in population 1 and 2 however magnified in population 3 below temperature (35 and 40°C) as compared with the control (Fig. 4a). Altogether populations, the SOD1 intensity was shrunken at 24 and 48 hrs with increase heat temperature (Fig. 4b, c), however, it was magnified at 10 days once heat temperature was (35 and 40°C) compared with heat-untreated seedlings (heat temperature 25°C) (Fig. 4d).
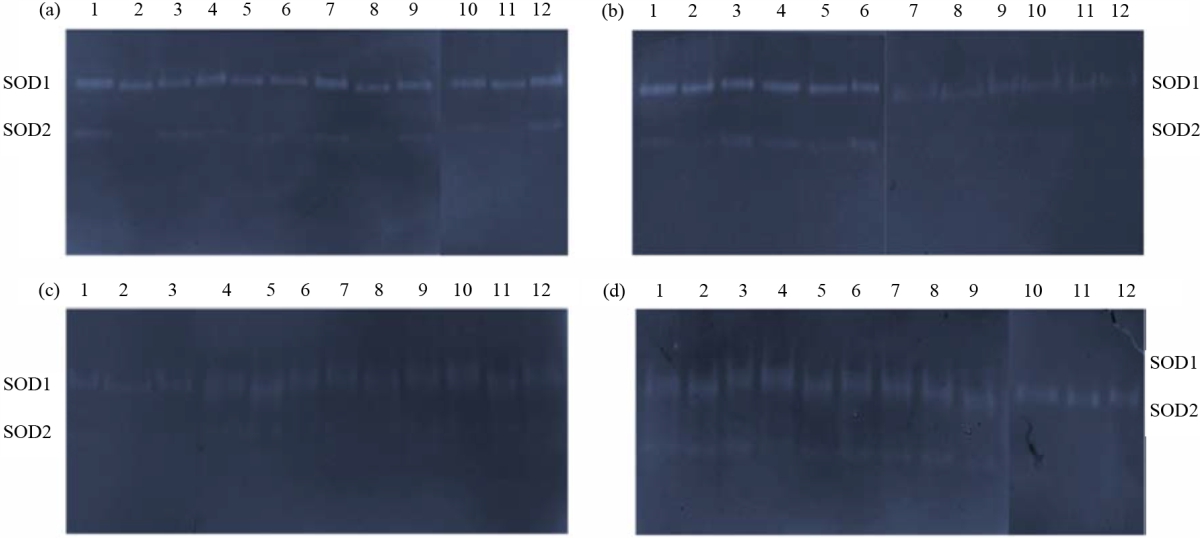 |
| Fig. 4(a-d): | Superoxide dismutase expression in seedlings of three populations of Tetraena propinqua under heat stress after exposure for (a) 4 hrs, (b) 24 hrs, (c) 48 hrs and (d) 10 days at each temperature 25°C (1, 2, 3); 30°C (4, 5, 6); 35°C (7, 8, 9); 40°C (10, 11, 12). Population 1: (1, 4, 7, 10), Population 2: (2, 5, 8, 11), Population 3: (3, 6, 9, 12) |
The intensity of SOD2 with (Rf 0.50) increased at 4 hrs in population 3 but decreased in population 1 and disappeared in population 2 once heat temperature reaches (35 and 40°C) in comparison with normal temperature (Fig. 4a). It magnified within the 3 populations at 24 hrs bit by bit at 30°C heat temperature then disappeared with increase heat temperature, however, it disappeared within the 3 populations for 48 hrs under heat-untreated and heat-treated seedlings (Fig. 4b, c). The SOD2 intensity magnified bit by bit at 10 days altogether populations but disappeared below 40°C as compared with the conventional temperature and in comparison with previous hours of seedlings like 4, 24 and 48 hrs (Fig. 4d).
DISCUSSION
The total soluble protein content elicited in population one and three beneath 35°C however reduced in population two and altogether populations beneath four 40°C for 4 hrs as compared with the conventional temperature. At 24 hrs of germination, it reduced bit by bit in population one beneath 40°C however elicited in population three beneath 35°C and population two and three beneath 40°C. Once 48 hrs of germination, it elicited altogether populations once the temperature was augmented to 40°C. Once ten days, the full soluble protein reduced in population two and three beneath 40°C, however, elicited in population one as compared to manage. Heat stress (35, 40°C) evoked total soluble protein in populations 1 and 2 however reduced in population 3 with increasing exposure time to ten days, showed that heat stress resulted in an increase of some soluble proteins and a decrease of others. This could be because of protein denaturation and inhibition of the protein synthesis at higher temperatures since the injury from hot temperature has typically been attributed to the denaturation of proteins22. The induction and reduction in total soluble proteins underneath heat stress as per the findings of Xiaozhi23 in alternative species, reported that a decrease in soluble protein content with increasing temperatures. Gulen and Eris24, reported that total protein content was decreased by heat stress. Similarly, slashed total soluble protein content in mulberry leaf was according to underneath heat stress25. Heat stress gives rise to an excessive increase in membrane fluidity, a confusion of protein function and turnover and metabolic imbalances in plants5. To abolish the detrimental effects of heat stress, plants have appointed multiple stress-tolerant plannings by changing the gene expression, protein synthesis and post-translational modification, which participate in the repetition of cellular homeostasis for plant survival under high temperature6. The plant cells interact with heat stress by stimulating the transcription of genes encoding heat shock proteins (Hsp), which are implicated in preventing or repairing the damage caused by elevated temperature and thus give increased thermotolerance7.
The results of protein profile reportable that some polypeptides were newly synthesized as 23 KD below 35°C in population 3 and population 2 below 40°C for 4 hrs, 28 KD in population 1 below 30°C and 20 KD altogether populations below 40°C for at 48 hrs, Some polypeptides degraded as 39 KD underneath (30-40°C) in population 2, underneath 40°C in population 3 and 25 KD in population 1 underneath 30°C, two severally for 4 hrs, however, for 24 hrs polypeptides of 83 and 68 KD degraded all told populations beneath heat temperature 35°C. The consumption of 83, 68 and 40 KD polypeptides were solely beneath normal temperature in population 2, 3 and all told populations once heated temperature than normal temperature for 48 hrs, respectively, however, the polypeptide consumption for 10 days was 40 KD beneath (25 and 40°C) and 30 KD beneath untreated and treated expect in population one beneath 40°C. The expression of most polypeptides diminished for 4 hours however induced for 24, 48 hrs and ten days with increase heat temperature to 40°C relative to their expression among the management seedlings. It is the modification in the expression profile of existing similarly as new proteins that play a vital role in modulating the defense of the plant beneath completely different abiotic stresses. Protein degradation may be related to heat injury throughout either gradual will increase in temperature26.
The expression of new proteins of various molecular weights 20, 23, 28 KD, could be playing the role in making tolerance against heat stress. The low-molecular-mass HSPs are the foremost extensive proteins elicited by heat stress in higher plants27. The small heat shock proteins (sHsps) involve proteins with molecular weights within the range of 15-42 KD28. The sHsps can bind to partly folded or denatured a protein that prevents irreversible unfolding or incorrect protein aggregation or binds to unfolded proteins and permits additional refolding by Hsp70/Hsp100 complexes29. The results were in agreement with previous studies indicating that protein synthesis positively correlates with stress tolerance and heat-tolerant plants maintain better protein synthesis and a lower protein degradation rate than heat-intolerant plants30,31. Heat stress enhances thermotolerance by facilitating protein synthesis has been well stimulating the excess production of reactive oxygen species (ROS) in plant cells ends up in the prevalence of oxidative stress9,32,33.
The 3 genotypes of T. propinqua ssp. migahidii exhibited completely different responses of SOD expression to heat stress. Within the current study, the expression of SOD1 (Rf 0.27) reduced at 4 hrs once the temperature inflated step by step to 40°C in genotype one, two however elicited in genotype three below (35 and 40°C) as compared with the management conditions. Altogether genotypes, the SOD1 expression was lower below 35 and 40°C for 24 and 48 hrs than for 10 days as compared with the conventional conditions. The expression of SOD2 (Rf 0.50) was induced in genotype 3 but reduced in genotype 1 and disappeared in genotype 2 once heat temperature reaches (35 and 40°C) for 4 hrs. It magnified within the genotypes at 24 hrs bit by bit at 30°C heat temperature then disappeared with increase heat temperature, however, it disappeared for 48 hrs under heat-untreated and heat-treated seedlings. The SOD2 intensity elicited bit by bit at 10 days altogether populations but disappeared below 40°C as compared with the conventional temperature and in comparison with previous hours of seedlings like 4, 24 and 48 hrs.
Heat reduced SOD1 and SOD2 were detected altogether genotypes, however, occurred higher for (10 days) than (4, 24 and 48 hrs) below (35 and 40°C), that may be concerned in scavenging ROS created throughout semi-permanent heat stress. Furthermore, SOD expression might be reduced after 4 hrs at 40°C in showed that this SOD isozyme was relatively sensitive to heat treatment. In agreement with this result, a study in Arabidopsis reported that heterologous expression of a heat-tolerant SOD-like activity led to induced tolerance to heat stress34. Sainz 35, reported that heat-induced degradation of chloroplast Cu/Zn superoxide dismutase as shown by reducing protein levels and isozyme-specific SOD activity of Lotus japonicas. The results were in agreement with previous studies indicating that superoxide dismutase (SOD) is considered to have essential roles in scavenging Reactive Oxygen Species (ROS) and protecting cells from oxidative damage in higher plants10,36,37. SOD, a member of the metalloenzyme family, induces the disharmony of O2– ion into hydrogen peroxide (H2O2) and oxygen (O2) molecules. Heat stress progresses, the SOD activity might decrease due to ROS-mediated injury to cells operate, together with lipid peroxidation of cell membranes. Environmental stresses might lead to high protein turnover, leading to the necessity for brand new SOD protein synthesis to take care of SOD levels spare for effective protection38. The expression of the SOD enzyme was higher in genotype one than in genotypes two and three by 10 days of heat stress, indicated that genotype one was the most tolerant. Associate degree isozyme SOD1 was induced beyond SOD2 throughout heat stress39. The increased SOD activity was observed in the leaves of grape (Vitis vinifera) cultivars resistant to lime-induced chlorosis compared with less resistant cultivars, it was considered a protective mechanism against the formation of superoxide40. SOD has been qualified to be implicated in several abiotic stress-related responses like heat, drought, salt, low-temperature conditions11-13.
The stress-specific HSPs expression was taken as a molecular system by that plants can tolerate heat stress. The expression of proteins associated with stress indicates that Tetraena propinqua ssp. migahidii heat adaptation in the early hours of germination and additional studies are required for its protein identification in the future.
CONCLUSION
The results showed that heat stress (35 and 40°C) elicited or reduced total soluble proteins, indicated that plants have appointed multiple stress-tolerant. The expression of 23, 28 and 20 KD indicated the upregulation of new HSPs proteins after heat stress. The overexpression of most proteins induced for 24, 48 hrs and 10 days with stress can enhance heat tolerance. SOD expression reduced in the first of exposure to heat stress and induced in the last stage, indicated a protective mechanism against the formation of superoxide. Heat stress indicated a negative relation between soluble protein and SOD expression.
SIGNIFICANCE STATEMENT
This study discovers the important aspect for heat acclimatization in total soluble protein and superoxide dismutase that can be beneficial for further sustainable protein identification techniques in the future. This study will help the researcher to uncover the critical areas of other protein and superoxide dismutase technologies that many researchers were not able to explore. Thus a new theory on advanced protein identification may be arrived at.
ACKNOWLEDGMENTS
This research was funded by Deanship of Scientific Research at Princess Nourah bint Abdulrahman University. (Grant No 39-S-268).
REFERENCES
- Baier, M. and K.J. Dietz, 2005. Chloroplasts as source and target of cellular redox regulation: A discussion on chloroplast redox signals in the context of plant physiology. J. Exp. Bot., 56: 1449-1462.
CrossRefDirect Link - D'Autreaux, B. and M.B. Toledano, 2007. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol., 8: 813-824.
CrossRefPubMedDirect Link - Schwarzländer, M. and I. Finkemeier, 2013. Mitochondrial energy and redox signaling in plants. Antioxid. Redox. Signal., 18: 2122-2144.
CrossRefDirect Link - Guan, Q., X. Lu, H. Zeng, Y. Zhang and J. Zhu, 2013. Heat stress induction of mi R 398 triggers a regulatory loop that is critical for thermotolerance in A rabidopsis. Plant J., 74: 840-851.
CrossRefDirect Link - Moreno, A.A. and A. Orellana, 2011. The physiological role of the unfolded protein response in plants. Biol. Res., 44: 75-80.
CrossRefDirect Link - Bokszczanin, K., 2013. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci., Vol. 4.
CrossRefDirect Link - Charng, Y.Y., H.C. Liu, N.Y. Liu, W.T. Chi, C.N. Wang, S.H. Chang and T.T. Wang, 2007. A heat-inducible transcription factor, HsfA2, is required for extension of acquired thermotolerance in Arabidopsis. Plant Physiol., 143: 251-262.
CrossRefDirect Link - Reddy, P.S., G. Mallikarjuna, T. Kaul, T. Chakradhar, R.N. Mishra, S.K. Sopory and M.K. Reddy, 2010. Molecular cloning and characterization of gene encoding for cytoplasmic Hsc70 from Pennisetum glaucum may play a protective role against abiotic stresses. Mol. Genet. Genomics, 283: 243-254.
CrossRefDirect Link - Almeselmani, M., P.S. Deshmukh, R.K. Sairam, S.R. Kushwaha and T.P. Singh, 2006. Protective role of antioxidant enzymes under high temperature stress. Plant Sci., 171: 382-388.
CrossRef - Gao, Y., Y.K. Guo, S.H. Lin, Y.Y. Fang and J.G. Bai, 2010. Hydrogen peroxide pretreatment alters the activity of antioxidant enzymes and protects chloroplast ultrastructure in heat-stressed cucumber leaves. Sci. Hortic., 126: 20-26.
CrossRefDirect Link - Wang, W., X. Zhang, F. Deng, R. Yuan and F. Shen, 2017. Genome-wide characterization and expression analyses of superoxide dismutase (SOD) genes in Gossypium hirsutum. BMC Genomics, Vol. 18.
CrossRefDirect Link - Tyagi, S., S. Sharma, M. Taneja, Shumayla, R. Kumar, J.K. Sembi and S.K. Upadhyay, 2017. Superoxide dismutases in bread wheat (Triticum aestivum L.): Comprehensive characterization and expression analysis during development and, biotic and abiotic stresses. Agri Gene, 6: 1-13.
CrossRefDirect Link - Feng, K., J. Yu, Y. Cheng, M. Ruan and R. Wang et al., 2016. The SOD gene family in tomato: Identification, phylogenetic relationships and expression patterns. Front. Plant Sci., Vol. 7.
CrossRefDirect Link - Beier, B.A., M.W. Chase and M. Thulin, 2003. Phylogenetic relationships and taxonomy of subfamily Zygophylloideae (Zygophyllaceae) based on molecular and morphological data. Plant Syst. Evol., 240: 11-39.
CrossRefDirect Link - Ghazanfar, S.A. and J. Osborne, 2015. Typification of Zygophyllum propinquum Decne. and Z. coccineum L. (Zygophyllaceae) and a key to tetraena in SW Asia. Kew Bull., Vol. 70.
CrossRefDirect Link - Alzahrani, D.A., 2019. Systematic studies on the Zygophyllaceae of Saudi Arabia: Two new subspecies combination in Tetraena maxim. Saudi J. Biol. Sci., 26: 57-65.
CrossRefDirect Link - Kruger, N.J., 1996. The Bradford Method For Protein Quantitation. In: The Protein Protocols Handbook: Springer Protocols Handbooks, Walker, J.M. (Ed.), Humana Press, Totowa, ISBN: 978-1-59745-198-7, pp: 15-20.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Kang, D.H., Y.S. Gho, M.K. Suh and C.H. Kang, 2002. Highly sensitive and fast protein detection with coomassie brilliant blue in sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Bull. Korean Chem. Soc., 23: 1511-1512.
CrossRefDirect Link - Chen, C.N. and S.M. Pan, 1996. Assay of superoxide dismutase activity by combining electrophoresis and densitometry. Bot. Bull. Acad. Sin., 37: 107-111.
Direct Link - Weydert, C.J. and J.J. Cullen, 2010. Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nat. Protoc., 5: 51-66.
CrossRefDirect Link - Xiaozhi, Z.R.F.Z.L. and W.Z.H. Wei, 1995. The effect of heat acclimation on membrane thermostability and relative enzyme activity. Acta Agron. Sinica, Vol. 5.
Direct Link - Gulen, H. and A. Eris, 2004. Effect of heat stress on peroxidase activity and total protein content in strawberry plants. Plant Sci., 166: 739-744.
CrossRefDirect Link - Chaitanya, K.V., D. Sundar and A.R. Reddy, 2001. Mulberry leaf metabolism under high temperature stress. Biol. Plant., 44: 379-384.
CrossRefDirect Link - He, Y., X. Liu and B. Huang, 2005. Protein changes in response to heat stress in acclimated and nonacclimated creeping bentgrass. J. Am. Soc. Hortic. Sci., 130: 521-526.
CrossRefDirect Link - Vierling, E., 1991. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: 579-620.
CrossRefDirect Link - Sun, W., M. Van Montagu and N. Verbruggen, 2002. Small heat shock proteins and stress tolerance in plants. Biochem. Biophys. Acta, 1577: 1-9.
CrossRef - Al-Whaibi, M.H., 2011. Plant heat-shock proteins: A mini review. J. King Saud Univ.-Sci., 23: 139-150.
CrossRefDirect Link - Pastenes, C. and P. Horton, 1996. Effect of high temperature on photosynthesis in beans (I. Oxygen evolution and chlorophyll fluorescence). Plant Physiol., 112: 1245-1251.
CrossRefDirect Link - Wang, L., C. Xu, C. Wang and Y. Wang, 2012. Characterization of a eukaryotic translation initiation factor 5A homolog from Tamarix androssowii involved in plant abiotic stress tolerance. BMC Plant Biol., Vol. 12.
CrossRefDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Wilson, R.A., M.K. Sangha, S.S. Banga, A.K. Atwal and S. Gupta, 2014. Heat stress tolerance in relation to oxidative stress and antioxidants in Brassica juncea. J. Environ. Biol., Vol. 35.
Direct Link - Im, Y.J., M. Ji, A. Lee, R. Killens, A.M. Grunden and W.F. Boss, 2009. Expression of Pyrococcus furiosus superoxide reductase in arabidopsis enhances heat tolerance. Plant Physiol., 151: 893-904.
CrossRefDirect Link - Sainz, M., P. Díaz, J. Monza and O. Borsani, 2010. Heat stress results in loss of chloroplast Cu/Zn superoxide dismutase and increased damage to photosystem II in combined drought-heat stressed Lotus japonicus. Physiol. Plant., 140: 46-56.
CrossRefDirect Link - Apel, K. and H. Hirt, 2004. Reactive oxygen species: Metabolism, oxidative stress and signal transduction. Annu. Rev. Plant Biol., 55: 373-399.
CrossRefPubMedDirect Link - Pnueli, L., H. Liang, M. Rozenberg and R. Mittler, 2003. Growth suppression, altered stomatal responses and augmented induction of heat shock proteins in cytosolic ascorbate peroxidase (Apx1)-deficient Arabidopsis plants. Plant J., 34: 187-203.
CrossRefDirect Link - Scandalios, J.G., 1993. Oxygen stress and superoxide dismutases. Plant Physiol., 101: 7-12.
CrossRefPubMedDirect Link - He, Y. and B. Huang, 2010. Differential responses to heat stress in activities and isozymes of four antioxidant enzymes for two cultivars of Kentucky bluegrass contrasting in heat tolerance. J. Amer. Soc. Hort. Sci., 135: 116-124.
CrossRefDirect Link - Ostrovskaya L.K., V.V. Truch and O.M. Mikhailik, 1990. Superoxide dismutase activation in response to lime-induced chlorosis. New Phytol., 114: 39-45.
CrossRefDirect Link