
Retno Sri Iswari
Department of Biology, Faculty of Mathematics and Natural Science, Universitas Negeri Semarang, Indonesia
LiveDNA: 62.35022
Muchamad Dafip
Department of Biology, Faculty of Mathematics and Natural Science, Universitas Negeri Semarang, Indonesia
LiveDNA: 62.35020
Eling Purwantoyo
Department of Biology, Faculty of Mathematics and Natural Science, Universitas Negeri Semarang, Indonesia
Pakistan Journal of Biological Sciences
Year: 2021 | Volume: 24 | Issue: 3 | Page No.: 319-325







ABSTRACT
Background and Objective: Malondialdehyde (MDA) may increase influenced by free radicals due to lipid oxidation. Tomato induction considers able to prevent free radical damage and atherosclerosis. Therefore, this study aims to understand the effect of steamed-tomato extracts on MDA and its potential as an early diagnosis of atherosclerosis. Materials and Methods: A total of 24 healthy 12 weeks-old male-rats were divided into four treatment groups, equally. A normal control group (K1) was rats with placebo treatment. A negative control group (K2) was the rats supplemented with 2 mL kg–1 b.wt. per day of cholesterol until cholesterol. A K3 group was atherosclerosis rats given with 20 mg kg–1 b.wt. per day of atorvastatin and a K4 was atherosclerotic rats supplemented with 16 mg kg–1 b.wt. per day of tomato extract. All treatments were carried out for 60 consecutive days. Results: Tomato extract in the K4 group was succeeded in lowering MDA production. Carotenoid compounds in tomato extract are well known to be prevention agents against lipid oxidation and inhibit free radicals. MDA levels have increased significantly in atherosclerosis conditions, making it potentially noticeable during early atherosclerotic, therefore, potentially developed as biomarkers. Conclusion: MDA levels increase significantly and simultaneously after high cholesterol diets and in line with lipid parameters and damaged blood vessels. The steamed-tomato extract can reduce MDA, lipids levels and protect endothelial from lipid oxidation. More research should be conducted to breakdown the MDA function in the molecular pathway, including MDA correlation to microRNA expression and cell signaling.
PDF Abstract XML References Citation
Received: December 02, 2020;
Accepted: February 06, 2021;
Published: February 15, 2021
Copyright: © 2021. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Retno Sri Iswari, Muchamad Dafip and Eling Purwantoyo, 2021. Malondialdehyde (MDA) Production in Atherosclerosis Supplemented with Steamed Tomato. Pakistan Journal of Biological Sciences, 24: 319-325.
DOI: 10.3923/pjbs.2021.319.325
URL: https://scialert.net/abstract/?doi=pjbs.2021.319.325
DOI: 10.3923/pjbs.2021.319.325
URL: https://scialert.net/abstract/?doi=pjbs.2021.319.325
INTRODUCTION
Atherosclerosis is a manifestation of lipid accumulation in blood vessels and oxidation by free radicals1,2. The majority of atherosclerosis patients cannot early diagnose because of no specific symptoms. Research is needed to find specific biomarkers in the early diagnosis of atherosclerosis3. The presence of chemical signaling compounds in oxidative stress may be useful as a biomarker on early detection of the disease development4. Malondialdehyde (MDA) is a biological marker produced mainly in high free-radical conditions and depicted an early cellular metabolic abnormality and disease development5,6.
Tomato extract is investigated and proved to reduce total cholesterol levels7,8, triglycerides, LDL-cholesterol and increases HDL-cholesterol levels9. The carotenoids in tomatoes are actively involved in activating the Retinoid A Receptor (RAR) and Retinoid X Receptor (RXR) of nuclear receptors, in decreasing inflammation10. The content of carotenoids such as β-carotene and lycopene, vitamins, including ascorbic acid and α-tocopherol, can block free radicals in triggering metabolic inflammation11,12. In previous research, the power of tomato’s nutrition and antioxidant activity can be increased and strengthen by steam-processing8.
Based on the description above, this study aims to determine the effect of the steamed-tomato extract on MDA concentrations and to evaluate the potential of MDA as a biomarker for early diagnosis of atherosclerosis. Identification of MDA and its correlation to cholesterol is an innovative, effective and efficient alternative to developing non-invasive biomarkers for early diagnosis of atherosclerosis.
MATERIALS AND METHODS
Study area: The research was conducted at the Biochemistry Laboratory of the Department of Biology, the Universitas Negeri Semarang, Indonesia and the Laboratory of Molecular Biology, Faculty of Medicine, Universitas Gadjah Mada, Indonesia. This research project was conducted from 5th March-30th August, 2020. Licensing for the use of experimental animals or Animal Care from the Ethics Commission, Faculty of Medicine, Public Health and Nursing, Gadjah Mada University, Indonesia registered number: KE/ FK/ 1003/ EC/ 2019.
Methodology: A total of 24 healthy 12-weeks-old male-rats with homogeneous body weight were divided into four treatment groups, with each group consisting of six rats. Each group was placed in a 2×1 meter cage fed with standard feed 594 pellets (PT Japfa Confeed Indonesia) ad libitum. The first group was normal to control (K1), which were normal rats treated with a placebo. The second group or negative control (K2) was rats supplemented with 2 mL of cholesterol every day until atherosclerosis. The third group or positive control group (K3), was contained atherosclerotic rats, which were given 20 mg kg–1 b.wt. of atorvastatin. The fourth group (K4) of atherosclerotic rats supplemented tomato extract at a dose of 16 mg kg–1 b.wt. All treatments were carried out for 60 consecutive days. This study used a posttest randomized controlled group design model
Tomato extraction: Tomato extract was obtained from 50 kg of thinly cut tomatoes, then steamed for 30 min at a temperature of 120°C8, then dried in the oven at 40-50°C for three days. After dried, the tomato was ground into a coarse powder and sieved using a sieve No. 100. The obtained fine powder was weighed as much as 50 g. Furthermore, the soxhlation process was carried out with petroleum ether as a solvent. The waste was dissolved with 500 mL of methanol as a solvent. Soxhletation was stopped until a clear filtered solution was obtained. The extract was mixed in a beaker glass. The obtained extract was then divided into Petri dishes and oven at 40-50°C to form a paste. The tomato extract was suspended in water for conditioning in mice13. Tomato lycopene extract was given once a day for 60 consecutive days.
Blood extraction and serum production: Blood collection was carried out every week through the retro-orbital plexus (corner of the eye) using a microhematocrit. The blood that comes out was collected in a 3 mL l tube-contained EDTA to obtain plasma. Blood plasma obtained from mice was measured total lipid consisting of High-Density Lipoprotein-Cholesterol (HDL-C), Low-Density Lipoprotein-Cholesterol (LDL-C), Triglyceride (TG) and free cholesterol, using the Diagnosis System (DiaSys) GmbH for metabolic syndrome marker (Holzheim, German), with spectrophotometric techniques (according to the manufacturer's instructions/ methods of action for each product). Each of 10-100 μL of serum was used in each test. The testing was carried out under the way it works and the manufacturer's protocol. Meanwhile, MDA was tested through plasma condensation and thiobarbituric acid and confirmed with 1-Methyl-2-phenylindole (as a more selective alternative reagent) and observed using spectrophotometry14.
Statistical analysis: The normality test for MDA and total lipid parameters was done by using the Shapiro-Wilk test. Then analyzed using One-way ANOVA with a confidence level of 95% and followed by LSD, with a confidence level of 95%. The analysis was performed using Statistical Product and Service Solution (SPSS) 24 for Windows data processing and presentation facilities.
RESULTS
The analysis showed that giving a high-cholesterol diet significantly increases total lipid levels in blood plasma (Table 1). The highest lipid levels have resulted in K2 rats, including triglycerides up to 171.35±3.41 mg dL–1, 214.78±6.00 mg dL–1 of total cholesterol and 84.38±1.99 mg dL–1 of LDL-C. Simultaneously, the lowest was in K1, there were 92.72±1.99 mg dL–1 of triglycerides, 108.92±2.72 mg dL–1 for total cholesterol and 39.56±1.56 mg dL–1 for LDL-C. The tomato extract administration (K4) was able to reduce the total lipid, including 107.43±4.24 mg dL–1 for triglyceride, 121.61±1.94 mg dL–1 for total cholesterol and 43.50±2.29 mg dL–1 for LDL-C levels significantly below K2 and K3. Besides, the high cholesterol diet induction increased the risk of developing atherosclerosis, as indicated by the atherogenic index value up to 0.79±0.04 or above 0.50.
Apart from total lipid levels, MDA levels also increased significantly from the 1st week of cholesterol induction to the last week of the study. Nonetheless, a decrease in MDA levels occurred in the K4 group at week four (Fig. 1a). A significant increase of MDA levels happened since the 5th week, which was 3.25±0.11 mmol mL–1 and increased sharply at the 7th week, reached 6.45±0.14 mmol mL–1. The average MDA on K2 increased by 52.43% every week until it reached a final concentration of 9.09±0.34 mmol mL–1 (Fig. 1b). The MDA level increases also occurred in all three groups, although at the 5th week, the MDA levels in K1, K3 and K4 were not as high as K2, which was 2.16±0.32 mmol mL–1 in K3, 1.83±0.10 mmol mL–1 in K4 and K1 which was 1.30±0.13 mmol mL–1. A significant decrease in the MDA level was observed in K4 after seven weeks of treatment, which showed 2.89±0.17 mmol mL–1. It then continued to decrease at the 8th and 9th weeks, there were 2.37±0.17 and 2.14±0.27 mmol mL–1, respectively. This was significantly different from K3, which increased from 2.65±0.28 mmol mL–1 in the 7th week and up to 3.51±0.30 mmol mL–1 and 3.55±0.26 mmol mL–1 in the 8th and 9th weeks.
| Table 1: Lipid parameters in each group after 60 days of treatment | |||||
| Parameter (mg dL-1) | K1 | K2 | K3 | K4 | p-value |
| Triglyceride | 92.72±1.99a | 171.35±3.41b | 120.00±2.45c | 107.43±4.24d | 0.000 |
| Total Cholesterols | 108.92±2.72a | 214.78±6.00b | 123.86±2.66c | 121.61±1.94c | 0.000 |
| LDL-C | 39.56±1.56a | 84.38±1.99b | 46.72±2.07c | 43.50±2.29d | 0.000 |
| HDL-C | 69.82±1.64a | 27.85±1.82b | 61.7±2.55c | 63.18±2.63c | 0.000 |
| VLDL-C | 42.15±0.91a | 77.88±1.55b | 54.55±1.12c | 48.83±1.93d | 0.000 |
| Atherogenic Index | 0.12±0.01a | 0.79±0.04b | 0.29±0.02c | 0.23±0.02d | 0.000 |
| AI risk criteria | Moderate | High | Moderate | Moderate | |
| LDL-C/HDL-C | 0.57±0.02a | 3.03±0.16b | 0.76±0.02c | 0.69±0.05d | 0.000 |
Different letters of the alphabet (a-d) show significant differences between treatment groups (p-value <0.05). An atherogenic index (AI) [Log (triglyceride / HDL-C)] value between -0.3-0.10 indicates a low cardiovascular risk, 0.10-0.24 indicates moderate risk and above 0.24 indicates high risk15,16. LDL-C: Low-density lipoprotein-cholesterol, VLDL-C: Very low-density lipoprotein-cholesterol, HDL-C: High-density lipoprotein-cholesterol | |||||
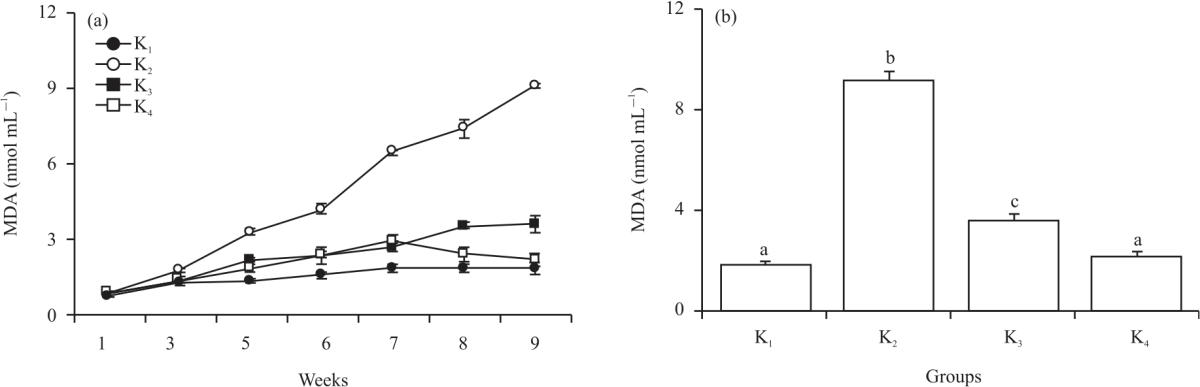 |
| Fig. 1(a-b): | Mean value of MDA levels every week (A) and the average MDA levels in the last week (B) Different letters of the alphabet (a-d) showed significant differences between treatment groups (p<0.05) |
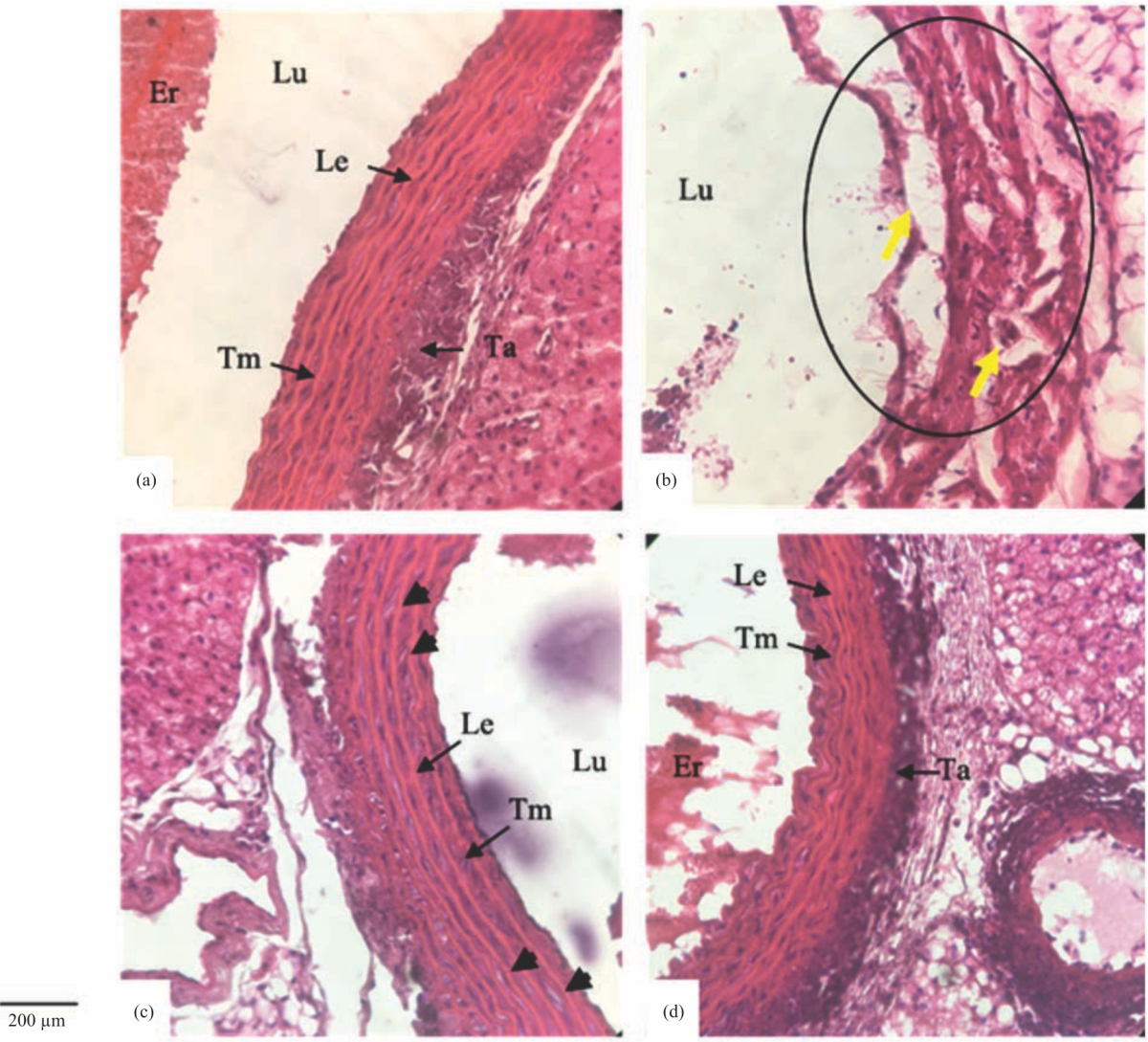 |
| Fig. 2(a-d): | Cross-section of the aorta of rats after 60 days of high cholesterol diet treatment in group, (a) K1, (b) K2, (c) K3 and K4 Intima damage in the form of fragmentation and deformation (B: dark ellipse); fatty smooth muscle cells (C: black arrowhead); and lesions (B: yellow arrows). Tm: Tunica media, Ta: Tunica adventitia, Le: Lamina elastica, Er: Erythrocytes, Lu: Lumens. Hematoxylin-Eosin (HE) staining, magnification and 400x microscope observation, 200 μm bar scale |
The reduction in MDA levels in the K4 group in the last week decreased so that it was not significantly different from K1 (Fig. 1b).
Cross-section of the aortic wall from the K1-K4 groups, showing the aortic wall condition after 60 days of treatment (Fig. 2a-d). In the cross-section, it can be identified the tunica media and tunica adventitia, except on K2. In the K1, K3 and K4 groups, the tunica media has a solid layer of lamina elastica and does not experience fragmentation. Besides that, the dark smooth muscle cell nucleus between the layers of the lamina elastica is observed.
Intima damage was observed in the K2 group of mice, characterized by total deformation of the tunica and elastic lamina (Fig. 2b). The damage is shown by the occurrence of fragmentation so that the lamina looks broken or not visible and the smooth muscle cells can no longer. Whereas in the K3 group, smooth muscle fat was observed in several parts, which were indicated by white spots (Fig. 2c). The shape of the blood vessel walls that are compact or solid and do not show any damage was observed in the rats’ blood vessel walls of K1 and K4 groups (Fig. 2a-d). This suggests that the tomato extract may minimize the occurrence of fat on the walls of blood vessels.
DISCUSSION
MDA levels in atherosclerotic rats rise with increasing intensity of cholesterol induction (consumption). The rise in MDA levels in atherosclerotic rats was in line with increased lipid parameters, including triglycerides, LDL-C and VLDL-C and decreased HDL levels. Also, in the atherosclerotic rats, lesion formation or endothelial damage was found in line with elevated blood MDA levels and high lipid parameters. The findings in this study indicate that MDA has the potential to identify or may a treatment target of atherosclerosis. These findings were not found in rats with a high cholesterol diet but also supplemented with 16 mg kg–1 b.wt. of steamed tomato extract. Administration of tomato extract showed inhibition of MDA productivity, total lipid parameters and prevented endothelial lesions
Increased synthesis of unsaturated fatty acids occurs due to the high input of cholesterol, which is degraded into acetic Ko-A and rearranged for storage as long-chain fatty acids (LCFA)17. This mechanism was evident in the K2 group, where the rats had increased levels of total blood lipids and fat in the blood vessel area. The high level of LCFA causes an increase in lipid oxidation, which increases the level of MDA. The MDA produces from the oxidation of unsaturated fatty acids, including arachidonic acid (C20:4), a-linolenic acid (C18:3) and linoleic acid (C18:2) as a byproduct of enzymatic processes. It is happened during the biosynthesis of thromboxane A2 (TXA2) and 12-l- hydroxy-5,8,10-heptadecatrienoic acid (HHT) or through a nonenzymatic process by cyclic endoperoxides produced during lipid peroxidation18. This compound is a reactive aldehyde group and is a reactive electrophile species that cause oxidative stress in cells. The rate of lipid peroxidation can be estimated by the amount of malondialdehyde in the tissue18.
High cholesterol consumption triggers LCFA biosynthesis while increasing the production of Radical Oxygen Species (ROS). Besides, cholesterol buildup causes atherosclerosis, which triggers endothelial inflammation1. As an aldehyde product, MDA can be used as a biomarker to measure the level of oxidative stress that occurs during the formation of atherosclerosis. This is evident in this study's results, where the increase in MDA levels is in line with the formation of atherosclerotic plaques on the walls of blood vessels and an increase in lipid levels.
This study also found that all parameters are not a significant increase in the group given tomato extract. In other words, giving tomato extracts can reduce MDA production. Three mechanisms are likely to contribute to the suppression of MDA production. The first mechanism involves the inhibition of tomato’s bioactive compounds, thereby reducing the production of endogenous cholesterol and using exogenous cholesterol to meet the body's physiological needs, so that body tissues do not accumulate cholesterol and fat7. This mechanism resembles the action of the drug atorvastatin, where the statins' chemical structure has a shape that resembles cholesterol and is a competitive inhibitor of the HMG-α reductase enzyme so that mevalonic acid is not formed from HMG-α19.
The second mechanism is that tomato extract acts as a regulatory gene. Tomatoes contain bioactive compounds, especially carotenoids, especially β-carotene and lycopene, undergo a metabolic process, especially in adipose cells20. Carotenoids are found in abundance in cells and adipose tissue in various forms but undergo conversion to retinoic acid as the main active form11. Molecular mechanisms mediate the effects of carotenoids on gene expression related to lipid biological processes in adipocytes.
Types of carotenoids from the carotene and provitamin A groups are used to synthesize retinoic acid, which has special receptors, namely Retinoid Acid Receptors (RARs) and Retinoid X Receptors (RXR) which are in the nucleoplasm membrane of all body cells. At least currently, three subtypes of RAR-RXR have been found, which include RARα, RARβ, RARγ and RXRα, RXRβ, RXRγ)10,11,21. The bond between the RA and the RAR or RXR complex will then bind the DNA sequence to the Retinoic Acid Response Element (RARE) or Retinoic X Response Element (RXRE) located in the promoter region of the retinoid target gene, either as a RAR-RXR or RXR-RXR dimmer. The carotenoids regulate gene expression in various cells through cell communication signaling pathways or via transcription factors involved in detoxification such as Aryl hydrocarbon Receptors (AhR), nuclear factor erythroid-2 linked factor 2 (NRF2) or PXR22,23. As an implication, the biological function of RA is supported by lycopenoic acid (an active compound derived from lycopene), which affects the liver through the Liver X Receptor (LXR), which increases HDL receptor synthesis and suppression of the synthesis of Apolipoprotein B (ApoB) and LDL24. The decrease in LDL synthesis results in a decrease in cholesterol distribution throughout the body, including the blood vessel network25.
In addition to regulating gene expression, the third mechanism is the bioactive content of tomatoes, especially carotenoids, which act as antioxidants that block ROS and at the same time act as immunomodulators for the body8. Carotenoid groups such as β-carotene bind various types of ROS, including singlet oxygen (1O2), superoxide anion radicals (•O2–), hydroxy radicals (OH–), peroxyl radicals (RCOO•) and nitric oxide (NO)26-28. The antioxidant activity of β-carotene and lycopene is influenced by the chemical structure in which the high amount of conjugated diene (covalent double bonds) will increase the scavenging activity of ROS, effectively.
Furthermore, ROS-type scavenger's effectiveness depends on the magnitude of the carotenoid Triplet Energy Transfer (TET) to the ROS's electron binding, a function of the conjugated double bond system (C=C)29,30. Lycopene is a carotenoid with an acyclic (free end) chain end model, which allows the reaction to occur by optimizing the TET of 11 conjugated double bonds to form polyenes. This is different from the cyclic carotenoid model, such as β-carotene, although it has the same number of double bonds as lycopene. The β-carotene chain end group's cyclization can reduce its efficiency as a ROS fastener due to loss of planarity of the two double bonds in the terminal or closed bond type (resulting in a large increase in TET)28.
Despite differences in ineffectiveness, the carotenoids in tomatoes can reduce ROS and give cells protection toward radical electrons. Besides, acyclic carotenes such as lycopene are considered the most robust carotenoid species in tomatoes because they accumulate stably at high concentrations. This causes lycopene to be more efficient in reducing ROS, especially 1O2, to reduce ROS significantly. Tomato carotenoids effectively break down lipid oxidation chain reactions with their ability to quell lipid peroxyl radicals29. The process of eliminating peroxyl radicals (ROO•) by lycopene and other carotenoids involves several mechanisms, namely: (a) electron transfer from carotene to ROO•, (b) hydrogen abstraction from ROO• to carotene and or, (c) incorporation of carotene with ROO•.
The regulation of gene expression and antioxidant mechanism presented by tomato extract to reduce the production of lipids and free radicals works simultaneously and influences each other. Various types of cell signaling, such as Mitogen-activated Protein Kinase (MAPK), Janus Kinase-Signal Transducer and Activator of Transcription (JAK-STAT) and necrosis factor kappa-beta (Nf-ĸB) pathway involve transcription of genes that affect cellular levels of ROS22. Furthermore, the presence of ROS directly can also increase or inhibit the various signaling pathways. Therefore, high levels of oxidative stress conditions that reflect the disease's pathophysiology can be investigated from high levels of MDA.
CONCLUSION
The MDA levels increase significantly and simultaneously after a high cholesterol diet and in line with the raised level of triglyceride, LDL-C, VLDL-C and damaged blood vessels. Furthermore, 16 mg kg–1 b.wt. of the steamed-tomato extract can reduce MDA, lipids levels and protect endothelial from lipid oxidation. Then, profiling of MDA and lipids parameters may give a holistic overview of the potential biomarker to identify atherosclerosis progression. More research should be conducted to breakdown MDA function in the molecular pathway to expand potential biomarkers for atherosclerosis detection, especially from transcriptomic subjects.
SIGNIFICANCE STATEMENT
This study discovers (1) the efficacious effect of steamed-tomato supplementation reducing lipid peroxidation on atherosclerosis progression, (2) the potential MDA as a potent biomarker for atherosclerosis profile diagnosis or possible treatment target. This study will help the clinical to develop potential alternative medicine for lipid metabolism disorder and for the researcher to uncover the critical areas of cellular mechanism in lipid peroxidation on atherosclerosis progression that many researchers were not able to explore. Thus, a new theory on early atherosclerosis diagnosis may be arrived at.
ACKNOWLEDGMENT
Acknowledgments are given to Universitas Negeri Semarang through the Institute for Research and Community Service for funding this research with contract number 232.23.4/UN37/ PPK.3.1/2020, April 23, 2020.
REFERENCES
- Conti, P. and Y. Shaik-Dasthagirisaeb, 2015. Atherosclerosis: A chronic inflammatory disease mediated by mast cells. Cent. Eur. J. Immunol., 3: 380-386.
CrossRefDirect Link - Abdolmaleki, F., S.M.G. Hayat, V. Bianconi, T.P. Johnston and A. Sahebkar, 2019. Atherosclerosis and immunity: A perspective. Trends Cardiovasc. Med., 29: 363-371.
CrossRefDirect Link - Rathouska, J., K. Jezkova, I. Nemeckova and P. Nachtigal, 2015. Soluble endoglin, hypercholesterolemia and endothelial dysfunction. Atherosclerosis, 243: 383-388.
CrossRefDirect Link - Madrigal-Matute, J., N. Rotllan, J.F. Aranda and C. Fernández-Hernando, 2013. MicroRNAs and atherosclerosis. Curr. Atheroscler. Rep., 15: 322-340.
CrossRefDirect Link - Roehrs, M., J. Valentini, C. Paniz, A. Moro and M. Charão et al., 2011. The relationships between exogenous and endogenous antioxidants with the lipid profile and oxidative damage in hemodialysis patients. BMC Nephrol., 12: 1-9.
CrossRefDirect Link - Rahmawati, I.S., S. Soetjipto, A.C. Adi and A. Aulanni'am, 2018. Malonaldehyde level of administration ethanol extract of purple sweet potato var. ayamurasaki in doca-salt hypertensive rats. J. Appl. Food Tech., 5: 6-9.
CrossRefDirect Link - Deplanque, X., D. Muscente-Paque and E. Chappuis, 2016. Proprietary tomato extract improves metabolic response to high-fat meal in healthy normal weight subjects. Food Nutr. Res., Vol. 60.
CrossRefDirect Link - Iswari, R.S. and R. Susanti, 2016. Antioxidant activity from various tomato processing. Biosaintifika: J. Biol. Biol. Educ.
CrossRefDirect Link - Mozos, I., D. Stoian, A. Caraba, C. Malainer, J.O. Horbańczuk6 and A.G. Atanasov, 2018. Lycopene and vascular health. Front. Pharmacol.
CrossRefDirect Link - Takeuchi, H., A. Yokota-Nakatsuma, Y. Ohoka, H. Kagechika, C. Kato, S.Y. Song and M. Iwata, 2013. Retinoid X receptor agonists modulate Foxp3+ regulatory T cell and Th17 cell differentiation with differential dependence on retinoic acid receptor activation. J. Immunol., 191: 3725-3733.
CrossRefPubMedDirect Link - Zapata-Gonzalez, F., F. Rueda, J. Petriz, P. Domingo, F. Villarroya, A. de Madariaga and J.C. Domingo, 2007. 9-cis-Retinoic Acid (9cRA), a Retinoid X Receptor (RXR) ligand, exerts immunosuppressive effects on dendritic cells by RXR-dependent activation: Inhibition of peroxisome proliferator-activated receptor γ blocks some of the 9cRA activities and precludes them to mature phenotype development. J. Immunol., 178: 6130-6139.
CrossRefDirect Link - Retno Sri Iswari, Ari Yuniastuti, Muchamad Dafip 2016. Tomato Extract as an Immunomodulator in Mice (Mus musculus) Infected with Plasmodium berghei Pak. J. Nutr., 15: 515-518.
CrossRefDirect Link - Tokac, M., G. Taner, S. Aydin, A.B. Ozkardes and H.Z. Dundar et al., 2013. Protective effects of curcumin against oxidative stress parameters and DNA damage in the livers and kidneys of rats with biliary obstruction. Food Chem. Toxicol., 61: 28-35.
CrossRefPubMedDirect Link - Anisimovich, P.V., T.B. Pochinok and E.V. Tokareva, 2017. Spectrophotometric determination of proteins in biological fluids. J. Anal. Chem., 72: 1212-1218.
CrossRefDirect Link - Cai, G., G. Shi, S. Xue and W. Lu, 2017. The atherogenic index of plasma is a strong and independent predictor for coronary artery disease in the Chinese Han population. Medicine, Vol. 96.
CrossRefDirect Link - Shen, S.W., Y. Lu, F. Li, C.J. Yang and Y.B. Feng et al., 2018. Atherogenic index of plasma is an effective index for estimating abdominal obesity. Lipids Health Dis., 17: 4-9.
CrossRefDirect Link - Sukhorukov, V.N., V.A. Khotina, Y.S. Chegodaev, E. Ivanova, I.A. Sobenin and A.N. Orekhov, 2020. Lipid metabolism in macrophages: Focus on atherosclerosis. Biomedicines, 8: 1-15.
CrossRefDirect Link - Całyniuk, B., E. Grochowska-Niedworok, K. Walkiewicz, S. Kawecka, E. Popiołek and E. Fatyga, 2016. Malondialdehyde (MDA)-product of lipid peroxidation as marker of homeostasis disorders and aging. Ann. Acad. Med. Siles., 70: 224-228.
CrossRefDirect Link - Fried, L.F., 2008. Effects of HMG-CoA reductase inhibitors (statins) on progression of kidney disease. Kidney Int., 74: 571-576.
CrossRefDirect Link - Mounien, L., F. Tourniaire and J.F. Landrier, 2019. Anti-obesity effect of carotenoids: Direct impact on adipose tissue and adipose tissue-driven indirect effects. Nutrients, 11: 1-14.
CrossRefDirect Link - Elias, M.D.B., F.L. Oliveira, F.C.R. Guma, R.B. Martucci, R. Borojevic and A.J. Teodoro, 2019. Lycopene inhibits hepatic stellate cell activation and modulates cellular lipid storage and signaling. Food Funct., 10: 1974-1984.
CrossRefDirect Link - Zhang, J., X. Wang, V. Vikash, Q. Ye, D. Wu, Y. Liu and W. Dong, 2016. ROS and ROS-mediated cellular signaling. Oxid. Med. Cell. Longev., 2016: 1-18.
CrossRefDirect Link - Barros, M.P., M.J. Rodrigo and L. Zacarias, 2018. Dietary carotenoid roles in redox homeostasis and human health. J. Agric. Food Chem., 66: 5733-5740.
CrossRefDirect Link - Bonet, M.L., J. Ribot and A. Palou, 2012. Lipid metabolism in mammalian tissues and its control by retinoic acid. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids, 1821: 177-189.
CrossRefDirect Link - Thies, F., L.M. Mills, S. Moir and L.F. Masson, 2017. Cardiovascular benefits of lycopene: Fantasy or reality? Proc. Nutr. Soc., 76: 122-129.
CrossRefDirect Link - Nishino, A., H. Yasui and T. Maoka, 2017. Reaction and scavenging mechanism of β-carotene and zeaxanthin with reactive oxygen species. J. Oleo Sci., 66: 77-84.
CrossRefDirect Link - Kawata, A., Y. Murakami, S. Suzuki and S. Fujisawa, 2018. Anti-inflammatory activity of β-carotene, lycopene and tri-n-butylborane, a scavenger of reactive oxygen species. In vivo (Brooklyn), 32: 255-264.
CrossRefDirect Link - Edge, R. and T.G. Truscott, 2018. Singlet oxygen and free radical reactions of retinoids and carotenoids-a review. Antioxidants, Vol. 7, No. 1.
CrossRefDirect Link - Pospíšil, P., A. Prasad and Marek Rác, 2019. Mechanism of the formation of electronically excited species by oxidative metabolic processes: Role of reactive oxygen species. Biomolecules, 9: 258-278.
CrossRefDirect Link - Qu, Z., 2020. Reactivities of singlet oxygen: Open-shell or closed-shell? Phys. Chem. Chem. Phys., 22: 13373-13377.
CrossRefDirect Link