
H.N. Malik
Not Available
S.I. Malik
Not Available
S.R. Chughtai
Not Available
H.I. Javed
Not Available
Asian Journal of Plant Sciences
Year: 2004 | Volume: 3 | Issue: 1 | Page No.: 6-10







ABSTRACT
The objective was to assess the heterotic relationship among nine maize inbreds derived from temperate, subtropical and tropical material. The nine inbreds and their diallel crosses excluding reciprocals were evaluated for eleven biometric traits viz., days to pollen shedding, plant height, leaves plant-1, leaf area, ears plant-1, ear weight, grain moisture at harvest, kernel rows ear-1, kernels row-1, 100-kernel weight and grain yield. High heterotic effects were observed for plant height, ears plant-1, ear weight, kernels row-1, 100-kernel weight and grain yield. Average MP and HP heterosis was 17.2 and 2.8%, respectively, for grain yield. Hybrid NCML-1082xNCML-1083 was the highest yielding with 82.6 and 74.5% heterosis relative to MP and HP, respectively. Subtropical and tropical heterotic groups were distinct for high grain yield in this study. Tropical lines also gave some reasonably good heterosis with sub-tropical material for other plant characters and also within its own origin (intra-origin) in certain combinations.
PDF Abstract XML References Citation
How to cite this article
H.N. Malik, S.I. Malik, S.R. Chughtai and H.I. Javed, 2004. Estimates of Heterosis among Temperate, Subtropical and Tropical Maize Germplasm. Asian Journal of Plant Sciences, 3: 6-10.
DOI: 10.3923/ajps.2004.6.10
URL: https://scialert.net/abstract/?doi=ajps.2004.6.10
DOI: 10.3923/ajps.2004.6.10
URL: https://scialert.net/abstract/?doi=ajps.2004.6.10
INTRODUCTION
Maize (Zea mays L.) is now the world’s leading cereal in terms of production and productivity[1]. It is a well known fact that one of the top breakthroughs in modern agriculture came with the discovery of heterosis, which may be defined as the difference between the hybrid and the mean of the two parents[2]. The expression of heterosis in plant crosses has been recognized for nearly 250 years[3] though the genetic basis of heterosis is not yet clearly understood. Heterosis is one of the most widely used terms in plant breeding and has made a dramatic impact on the development of breeding methods and high yields for many field crops, vegetables, ornamentals and tree species during the 20th century[4]. Exploiting heterosis could dramatically reduce production costs by increasing yield levels while enhancing input use efficiency[5]. This phenomenon is one of the primary reasons for success of commercial maize industry as well as the success of plant breeding endeavors[6].
The introgression of exotic germplasm could increase the heterosis among maize populations[7]. Extensive work have been done to explore the effect of heterosis[8-12] with almost similar objective of finding superior combinations which yield higher and suit well in the respective environments. Soengas et al.[13] in their study, defined certain heterotic patterns within flint germplasm adapted to temperate conditions and found that the crosses yielded 30% more than parental cultivars. Vacaro et al.[14] found that additive effects were predominant suggesting that environment also influenced the manifestation of heterosis. Saleh et al.[15] evaluated a series of tropical maize hybrids and found that hybrids varied significantly for grain yield and most other traits under study.
Diallel crosses have been widely used in genetic research to investigate the inheritance of important traits among a set of genotypes[16]. They were devised specifically to investigate combining ability of the parental lines for the purpose of identification of superior parents for use in hybrid development program. This diallel study was made with the objective to find the heterotic relationship among temperate, subtropical and tropical material and evaluate their breeding potential in the prevailing environment.
MATERIALS AND METHODS
Nine maize inbreds were used as parents: QPM-1, QPM-3 and QPM-5 from temperate, NCML-1061, NCML-1071 and NCML-1078 from subtropical and NCML-1082, NCML-1083 and NCML-1084 from tropical material from the collection maintained by the Maize, Sorghum and Millet Programme, NARC, Islamabad. The crosses were made in a diallel fashion excluding reciprocals. Each ear was obtained by cross-fertilization to one tassel only. All the 36 hybrids and 9 parents were entries in the diallel. Hybrids along with their parental lines were grown in the next growing season in a randomized complete block design with two replications. Hills were overplanted and thinned after emergence for the final population density approximated 56,000 plants ha-1. The experimental unit was one row for each entry, 5 m long and 75 m apart and the final plant stand was 21 plants row-1.
Uniform cultural practices were accomplished in both replicates. The following plant traits were recorded before harvest: days to pollen shedding, plant height (from soil to the lowest tassel branch), number of leaves plant-1, leaf area and number of ears plant-1. After all the entries had completed dry matter accumulation, the ears were harvested, husked and weighed. Data for several traits like ear weight, grain moisture at harvest, number of kernel rows ear-1 and number of kernels row-1 were recorded. Based on gross total plot yield, grain yield plot-1 was calculated. 100-kernel weight was recorded after ears were dried for about two months to a constant moisture level of 15%.
Analysis of variance were completed for all the traits under investigation while plot means were compared using the LSD test. Both mid-parent (MP) and high parent heterosis (HPH) were estimated as percent increase (+) or decrease (–) of F1 hybrids over the respective mid-parents and high parents for all crosses.
RESULTS AND DISCUSSION
Analysis of variance revealed highly significant differences among means for all traits under investigation (P=0.01) except number of kernels row-1 where difference was significant only (P=0.05). For the number of leaves plant-1 and 100-kernel weight the difference was non significant (Table 1).
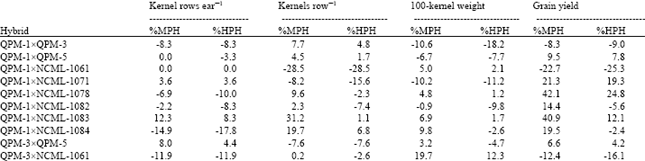
All the traits responded differently in the expression of heterosis in their respective combinations (Table 2). The average heterotic effect for days to pollen shedding remained negative which can be useful for the development of early maturing hybrids. High heterotic effects were observed in certain hybrids regarding plant height, especially lines from subtropical material gave superior combinations with MPH ranging from 13.6 to 37.8% and HPH ranging from 9.9 to 33.9% when crossed with lines from tropical material (NCML-1082, NCML-1083 and NCML-1084). Average MPH (8.4%) and HPH (1.1%) was moderate for this trait (Table 2).
Findings of this study revealed certain combinations with high heterotic effects for leaf area, although average MP and HP heterosis remained low (5.7 and -2.8, respectively). Some combinations (QPM-5xNCML-1082, NCML-1071xNCML-1082 and NCML-1078xNCML-1082) yielded higher MP and HP values (38.3 and 18.5, 33.4 and 23.6, 32.3 and 24.9%), respectively and were identified as the prominent heterotic groups for greater leaf area (Table 2). Average values of MPH (9.5%) and HPH (0.9%) were obtained for the number of ears plant-1 while moderate to high heterosis was observed in certain combinations (Table 2). Daud[17] reported high estimates of heterosis for number of ears plant-1 in their study. Hybrid QPM-1xNCML-1071 gave the highest values of 98.4 and 79.2% for MP and HP heterosis, followed by QPM-1xQPM-3 and NCML-1078xNCML-1083 with 58.2, 38.5% and 50.2, 43.9% MP and HP heterosis, respectively (Table 2).
Likewise, high heterotic effects were seen for ear weight in certain combinations (QPM-3xNCML-1082, NCML-1071xNCML-1084 and NCML-1082xNCML-1084) with MPH of 58.6, 58.4 and 53.4%, respectively (Table 2). HPH was found to be 47.0, 49 and 45.2% for the same hybrids. Grain moisture at harvest showed low heterosis (Table 2) which is a good sign towards high grain yield as grain moisture is negatively correlated with grain yield[18]. Revilla et al.[9] also reported insignificant heterotic effects for grain moisture at harvest in their study. The heterosis observed for kernel rows ear-1 was not significant. Moderate heterotic effects have been reported by Saleh et al.[15] whereas, significant by Revilla et al.[21] for kernels row-1. The highest MP and HP heterosis (25.9 and 21.4%) was observed for cross NCML-1071xNCML-1083 (Table 2).
Hybrids yielding greater number of kernels row-1 were NCML-1082xNCML-1083 (49.6 and 24.6% MP and HP heterosis), NCML-1078xNCML-1083 (42.4 and 20.3% MP and HP heterosis), followed by NCML-1071xNCML-1083 where MP and HP heterosis was 34.4 and 10.6%, respectively. Average MPH was 11.6% while HPH remained 1.5% (Table 2). Average heterosis relative to MP and HP for 100-kernel weight was 6.6 and 0.8%, respectively (Table 2), with the highest values of 34.5 and 22.5 for hybrids NCML-1061xNCML-1084, followed by 30.6 and 26.2% for QPM-3xNCML-1084 and, 29.0 and 16.2% for NCML-1071xNCML-1082.
High estimates of heterosis were observed for grain yield in this study (Table 2). The findings are in close agreement with those by Ordas[19], Azar et al.[11] and Saleh et al.[15]. Line NCML-1082 from tropical material was the highest yielding in combination with NCML-1083 from the same origin. This intra-origin heterotic group was identified as the best combination with 82.6 MP and 74.5% HP heterosis. The second high yielding hybrid was NCML-1078xNCML-1083 with 53.3% MP and 39.2% HP heterosis. Hybrids QPM-3xNCML-1078, NCML-1071xNCML-1084 and NCML-1083xNCML-1084 also gave high MP and HP heterosis of 55.2, 37.4%, 51.6, 25.5% and 43.0, 38.2%, respectively (Table 2).
| Table 1: | Mean squares from the analysis of variance, grand means and LSD for eleven traits for a diallel of nine maize inbreds |
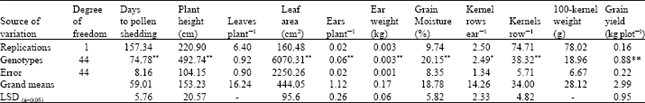 | |
| ** Significant at P=0.01, * Significant at P=0.05 | |
| Table 2: | Percent mid-parent heterosis (%MPH) and percent high parent heterosis (%HPH) for all the hybrids in their respective combinations |
 | |
 | |
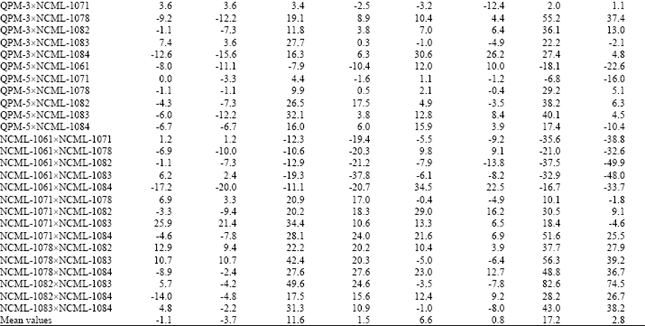 | |
Average heterosis for grain yield was 17.2% for MP and 2.8% for HP. Aguerre et al.[10] found grain yield MPH as 19%, Revilla and Tracy[12] reported it as 29.17% and 57.04% for two different planting dates while Soengas et al.[13] reported values ranging from 2.5 to 64.7%.
Gains in yield and yield stability offered by heterosis have prompted use of hybrids in several crops. The presence or absence of heterotic effects does not rule out the possibility that particular chromosomal segments carry either favorable or unfavorable alleles for a particular quantitative trait[20]. If the effects of alleles were additive, then a significant heterotic effect would not be expected. Additive gene action could be detected by a hybrid being significantly larger than one parent and significantly smaller than the other parent[20]. It is also commonly accepted that greater heterotic response is associated with greater genetic diversity of the parents[19]. In this study, tropical material was found to give some reasonably good heterosis for certain plant characters with sub-tropical material and within its own origin (intra-origin) in certain combinations. These findings have been very helpful in determining the two heterotic groups (subtropical and tropical) and also identifying intra-group combinations which, ultimately, could lead towards indigenous hybrid maize varieties with high grain yield.
REFERENCES
- Bertoia, L.M., R. Burak and M. Torrecillas, 2002. Identifying inbred lines capable of improving ear and stover yield and quality of superior maize hybrids. Crop Sci., 42: 365-372.
Direct Link - Azar, C., D.E. Mather and R.I. Hamilton, 1997. Heterosis and endosperm texture in crosses involving maize landraces of the St. Lawrence Great Lakes region of North America. Maydica, 42: 53-57.
CrossRef - Revilla, P. and W.F. Tracy, 1997. Heterotic pattern among open-pollinated sweet corn cultivars. J. Am. Soc. Hort. Sci., 122: 319-324.
Direct Link - Soengas, P., B. Ordas, R.A. Malvar, P. Revilla and A. Ordas, 2003. Heterotic pattern among flint maize populations. Crop Sci., 43: 844-849.
Direct Link - Vacaro, E., J.F.B. Neto, D.G. Pegoraro, C.N. Nuss and L.D.H. Conceicao, 2002. Combining ability of twelve maize populations. Pesq. Agropec. Bras. Brasilia, 37: 67-72.
CrossRefDirect Link - Saleh, G.B., D. Abdullah and A.R. Anuar, 2002. Performance, heterosis and heritability in selected tropical maize single, double and three-way cross hybrids. J. Agric. Sci., 138: 21-28.
CrossRefDirect Link - Lamkey, K.R., A.R. Hallauer and D.S. Robertson, 1988. Contribution of long arm of chromosome 10 to the total heterosis observed in five maize hybrids. Crop Sci., 28: 896-901.
Direct Link - Revilla, P., R.A. Malvar, C.P. Soengas and A. Ordas, 2002. Heterotic relationship among European maize inbreds. Euphytica, 126: 259-264.
CrossRef