
Rosemary I. Egonmwan
Department of Zoology, University of Lagos, Akoka, Lagos, Nigeria
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 1 | Page No.: 163-166







ABSTRACT
Rate of weight loss and mortality, of the Nigerian periwinkle, Tympanotonus fuscatus var. radula L. were investigated during prolonged exposure to air for one month. Short term thermal tolerance in the laboratory was carried out by heating the water in which the animals are placed at the rate of 1°C every 10 min. The group exposed to air continued to lose weight; when the experiment was terminated after 4 weeks, 20% of body weight was lost and 5 out of 40 periwinkles died, while the control experimental snails lost 11% of body weight, due to starvation. The species has an upper lethal temperature of 46-48.5°C, much higher than the temperature of its habitat. Survival of T. fuscatus var. radula out of water and adaptation to high temperature suggests the presence of highly adaptive mechanisms of water balance and metabolism.
PDF Abstract XML References Citation
How to cite this article
Rosemary I. Egonmwan, 2007. Thermal Tolerance and Evaporative Water Loss of the Mangrove Prosobranch Tympanotonus fuscatus var. radula L. (Cerithiacea: Potamididae). Pakistan Journal of Biological Sciences, 10: 163-166.
DOI: 10.3923/pjbs.2007.163.166
URL: https://scialert.net/abstract/?doi=pjbs.2007.163.166
DOI: 10.3923/pjbs.2007.163.166
URL: https://scialert.net/abstract/?doi=pjbs.2007.163.166
INTRODUCTION
Tympanotonus fuscatus var. radula L. commonly referred to in Nigeria as periwinkle though not a litorrinid snail is collected and sold for food in many coastal and urban cities of Nigeria. The collection and marketing of these snails form an important shell fish industry in the Niger Delta. Like many other edible species of molluscs, e.g., clams, mussels and oysters, the species is not cultivated in Nigeria: they are merely collected from natural beds at low tide. Though there is heavy pressure on periwinkle harvesting, stocks are not yet depleted, because the sizes harvested are the big sizes desired by the consumers, which may have spawned and some stock are located within parts of the mangrove swamp not accessible to the fishermen.
Tympanotonus fuscatus var. radula inhabits the mudflat of the Lagos lagoon system, which is a large expanse of shallow water, extending from Republic of Benin in the west to the Niger Delta in the east (Hill and Webb, 1958; Olaniyan, 1961; Yoloye, 1969; Fagade, 1969). The shores of the Lagos lagoon and adjacent creaks are lined in undeveloped areas by mangrove swamps. Although human activities such as the cutting of the mangrove tree, Rhizophora racemosa for firewood has altered the vegetation in many places, the fauna inhabiting the mudflats have remained typical of West Africa mangrove swamps. The habitat periodically dries up between November and April due to dry season and at low tides (Longhurst, 1958), exposing the animals. During the rainy season, the whole mudflat area is flooded by inflow of lagoon water but during the dry season some area may dry up leaving irregular anastomosing water channels. T. fuscatus var. radula, occurs in the mudflats of the mangrove swamp which are exposed except during high tides and during the rainy season when the area dries up. In the habitat, T. fuscatus var. radula, migrates to the water edge and congregates under tufts of grasses and breathing roots of the mangrove plants Avicenia nitida which shade them from the direct rays of the sun and during the dry season, when the mudflat dries up, the periwinkles burrow into the mud. The habitat in the Niger Delta is brackish all year round, never dries up and is flooded daily at high tide.
Millions of Tympanotonus species are transported from the Niger Delta daily to other parts of Nigeria in jute bags. They can remain in these bags for more than four weeks without water. During the transportation to the markets the periwinkles are subjected to high temperatures in addition to lack of food and water. Thermal tolerance has been used to examine the thermal capacities of marine invertebrates to tolerate their environment (Clarke et al., 2000), but in this study physiological tolerance is used to explain the survival of the species during the long period of transported to the market for sale. Eulittoral gastropods which are subjected to periodic but predictable rehydration have evolved adaptations that enable them to maintain their foraging activity during the short period of exposure whereas the eulittoral fringe species that experience prolonged and unpredictable emergence have selected for adaptations that minimize water loss rates (McMahon, 1990).
This study was undertaken in order to determine the effects of desiccation and temperature on the survival of these prosobranchs. The different rates of temperature changes and long period of exposure to air were designed to imitate the conditions that the periwinkles are exposed to during their transportation from the Niger Delta to the markets in Nigeria.
MATERIALS AND METHODS
These experiments were designed to imitating the conditions the periwinkles are subjected to during their transportation from their major habitat in the Niger Delta, by the traders, to markets all over Nigeria. To investigate the survival of T. fuscatus var. radula to dry conditions, 2 dry glass tanks were set up. To one tank (control), 40 weighed specimens of T. fuscatus var. radula were placed. To this tank filtered lagoon water (15.6 psu) to a height of 10 cm was added so that the animals were completely immersed. This aerated water was changed daily until the end of the experiment. Another 40 specimens of T. fuscatus var. radula were placed in the 2nd tank (experimental) but no water was added. Both tanks were kept on the laboratory table close to a window and the snails were not fed through out the duration of the experiment. The animals were weighed singly once a week and observations were made daily on their behaviour. The experiment was terminated at the end of the 4th week; the experimental snails were immersed in lagoon water to ascertain the rate of recovery.
To determine the upper thermal tolerance limit of T. fuscatus var. radula, heat coma treatment was carried out using a modification of the method of McMahon (1990). Adult specimens of T. fuscatus var. radula were collected at low tide and taken to the laboratory immediately after collection to acclimate to the laboratory temperature (28°C; l:d, 12:12). The specimens were measured to the nearest 0.1 mm with a Vernier caliper and 36 individuals of size range 55-60 mm shell length were selected for the experiment. The specimens were divided into 6 groups of 6 and each group was placed in a 200 mL beaker containing 100 mL of filtered lagoon water from the collection site (salinity, 15.6 psu) acclimated to laboratory to laboratory temperature of 28°C. The snails were allowed to move about the beaker and any inactive individual was replaced. The beaker was placed on 1 L beaker containing 500 mL of the lagoon water. The larger beaker was heated in a thermostat controlled water bath. The temperature of the water bath at the start of the experiment was the acclimated laboratory temperature of 28°C which was raised at the rate of 1°C every 10 min. A thermometer was placed into the beaker to monitor the temperature of water in it. At every degree increase, the operculum of the periwinkles was stimulated with a glass probe to ascertain if they were alive. The number of individuals displaying symptoms of heat coma in each beaker was recorded for every 1°C increase in temperature. When the animal showed no response to probing, it was removed from the hot water allowed to cool to room temperature to see if it would recover. The beakers were removed from the water both and the hot water contained in them poured away. The periwinkles were allowed to cool to room temperature, observed at 1 and 12 h post-heat treatment, the number of active and non active individuals recorded.
Temperature of the water, air and substratum of the habitat where the animals were collected were recorded on the day of collection.
RESULTS
At the end of the 1st day, all the periwinkles in the control experiment kept wet in tanks containing lagoon water became active. They were observed crawling about the bottom of the tank. The specimens in the dry tank were active on the first day, defecated and then withdrew into the shell. At the end of the 1st day only 5 of the experimental snails were protruded out of their shell. At the end of the 1st week, all the specimens in the dry tank were all withdrawn into their shells but were still alive. The specimens in the control experiment were all active, although they lost weight (Fig. 1). Experimental snails also lost weight after the 1st week, but more pronounced weight loss was high at 14% of body weight after the 1st week. The group continued to lose weight until the experiment was terminated at the end of week 4, when 20% of body weight was lost. The weight lost by the control experiment was 4% of body weight in the 1st week, 11% of body weight at the end of 4 weeks. When the experimental snails were immerse in lagoon water at the end of 4 weeks, all the snails became active, 5 were dead.
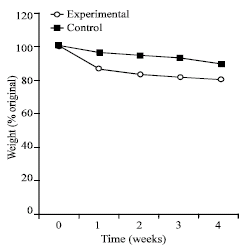 | |
| Fig. 1: | Percentage weight loss by T. fuscatus var. radula kept out of water and exposed to air. The experiment was terminated after 4 weeks of exposure |
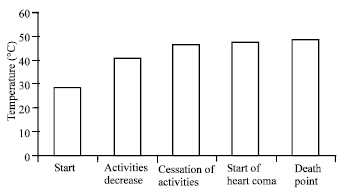 | |
| Fig. 2: | Distribution of temperature at which activities decrease and cease, upper thermal limit and death point of T. fuscatus var. radula when exposed to temperature increasing at the rate of 1°C 10 min-1 in the laboratory |
The result of the high temperature tolerance is shown in Fig. 2. At the beginning of the experiment, the periwinkles in the experimental group were active, their activity decreasing as the temperature of the water surrounding them approached 40°C. At 46°C, there was cessation of activities in all the animals; when the individuals were probed, they did not immediately withdraw into their shells as their movement was very sluggish. The start of heat coma was at 47°C in all the snails, when the periwinkles appeared dead but recovered after 1 h when they were taken out of the hot water. The death point of the periwinkle was 48.5°C, when they did not recover in lagoon water at 24°C.
The temperature for the mud flat was 29.5 and 30°C at the two times animals were collected for the experiment.
DISCUSSION
The absolute tolerance to desiccation probably determines the upper limits of vertical range for most intertidal species and adaptations by intertidal gastropods involves adjustments of several important physiological functions such as water economy, temperature resistance and metabolic resistant (Sokolova and Portner, 2001), for example increased resistance to prolonged air exposure and desiccation stress, is associated with improved water conservation abilities in Littorina saxatilis. T. fuscatus var. radula, during this study was withdrawn into the shell from the 1st week of the observation, probably to reduce evaporative water loss which may have been responsible for the small difference in body water loss between the experimental and control snails observed in this study. Tight closure of the operculum reduces evaporative water loss (Sokolova and Portner, 2001), but also reduce oxygen supply to tissues. A major intertidal gastropod adaptation is reduction of ctenidium surface area and formation of a mantle cavity lung (Fretter and Graham, 1976; McMahon, 1988); this may well be the case in T. fuscatus var. radula. The loss in the experimental snails could be attributed to starvation as food was not provided to either group through out the duration of this experiment.
The results of the thermal tolerance experiment clearly show that Tympanotonus fuscatus var. radula L. has a very high upper thermal limit so can tolerate very high temperatures. The periwinkles were active at very high temperature, the cessation of activities occurred at 46°C and heat coma started at 47°C when the periwinkles appeared dead but recovered after 1 h when they were taken out of the hot water. The death point of the periwinkle was 48.5°C, when they did not recover in lagoon water at 24°C. Heat coma temperature in gastropods, a reversible condition, is the temperature at which normal nervous function is lost, manifested by cessation of locomotion and inability to remain attached to substrata (McMahon, 1990). A mean coma tolerance of 31 to 41°C was reported for various species of intertidal prosobranchs, correlated with their habitat (McMahon, 1990); desiccation stress is reduced in individuals inhabiting below the high tide mark and increased in those above the high tide mark, therefore, the adaptation of the eulittoral species include evaporative cooling, low thermal tolerance and high capacity for radiant heat absorption in order to reduce evaporative water loss, while species limited to the eulittoral fringes show adaptations such as foot withdrawal, aestivation in air, elevated thermal tolerance and increased capacity for heat dissipation. The temperature in the mud flat is high all year round and exceptionally high during the dry season when the whole area dries up and the sun heats the soil directly. The temperature of the mud flat was about 30°C during the period of observation, the result of this study show that T. fuscatus var. radula appear to possess a higher capacity for thermal tolerance than some intertidal prosobranchs (Clark et al., 2000) was recorded in March 2005, therefore the T. fuscatus var. radula is likely to behave like the eulittoral fringe prosobranchs described earlier.
Therefore, the survival of T. fuscatus var. radula out of water suggests the presence of highly adaptive mechanisms of water balance and metabolism which makes them ideally adapted to life in the intertidal mangrove mudflats of the Lagos lagoon. This same tolerance is responsible for their ability to survive the harsh conditions they are subjected to during their transportation to markets all over Nigeria.
REFERENCES
- Clarke, A.P., P.J. Mill, J. Grahame and R.F. McMahon, 2000. Geographical variation in heat coma temperatures in Littorina species. J. Ma. Biol. Assoc. UK., 80: 855-863.
Direct Link - Hill, M.B. and J.E. Webb, 1958. The ecology of the Lagos lagoon. Part 11: The topography and physical features of the Lagos harbour and Lagos lagoon. Phil. Trans. R. Soc. Lond., B, 241: 319-333.
Direct Link - McMahon, R.F., 1990. Thermal tolerance, evaporative water loss, air-water oxygen consumption and zonation of intertidal prosobranchs: A new synthesis. Hydrobiologia, 193: 241-260.
Direct Link - Sokolova, I.M. and H.O. Portner, 2001. Physiological adaptations to high intertidal life involve improved water conservation abilities and metabolic rate depression in Littorina saxatilis. Mar. Ecol. Prog. Ser., 224: 171-186.
Direct Link